Training Your Tongue
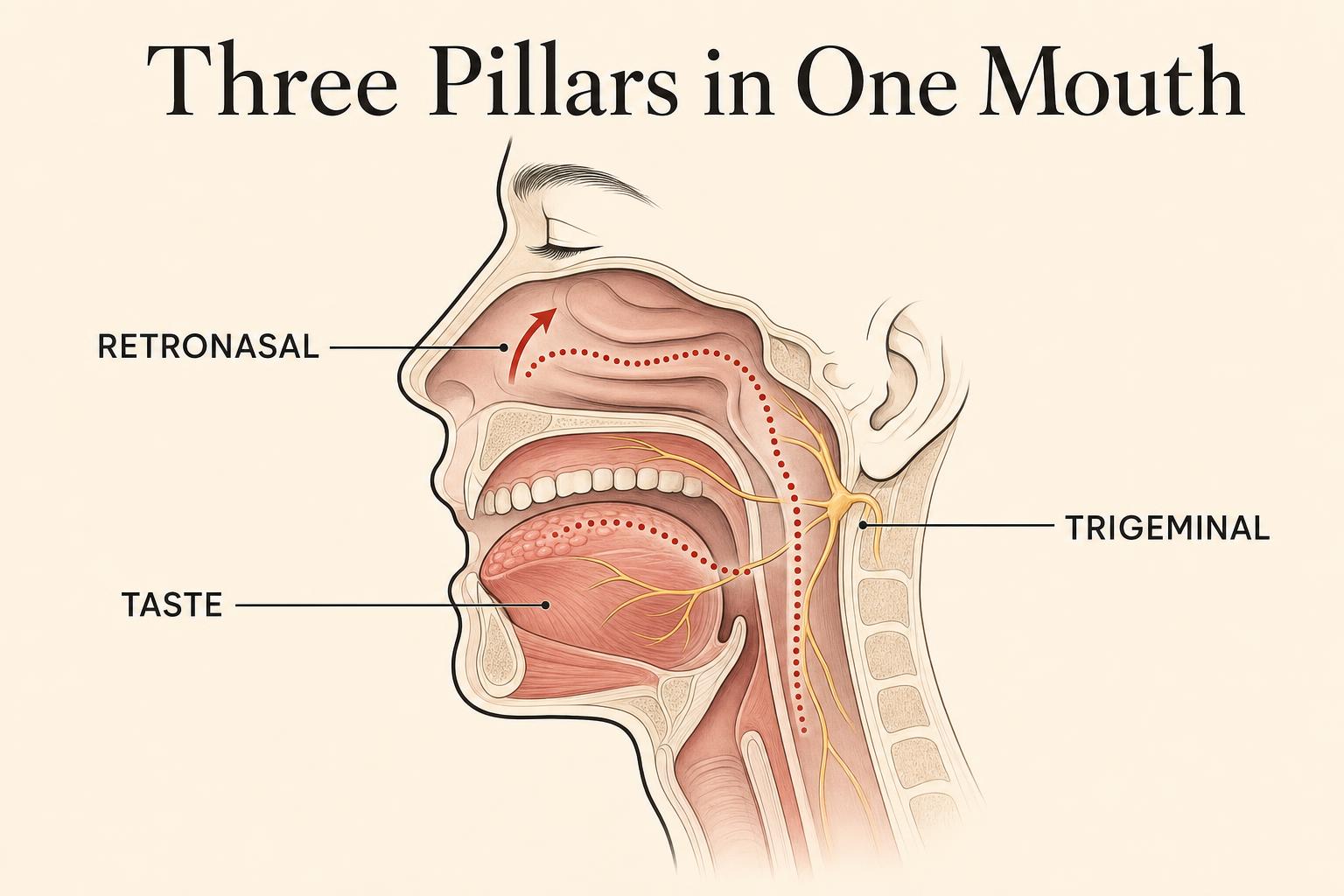
"Flavor" is not a single sense but a construction from three pillars — taste, retronasal smell, and the trigeminal nerve. All three can be trained.
Imagine two friends sharing a pot of aged sheng pu-erh. One leans back and sighs: "It's like walking through a wet forest after rain — mushrooms, camphor, a sweet mineral finish." The other frowns: "All I get is bitter and musty." They are drinking the same tea, brewed identically, poured from the same pot. Neither is wrong. The liquid in their cups is chemically identical, but the nervous systems receiving that liquid are not — and the languages those nervous systems have been trained to use are worlds apart.
This chapter is about the space between the cup and the experience. You have traced tea from mountain terroir through withering, oxidation, and firing. Now we follow those hard-won chemical compounds on their final journey: across the tongue, up through the nasal passages, and into the brain. Along the way, you will discover that what you casually call "flavor" is actually a symphony performed by three distinct sensory systems, shaped by your genes, your culture, and your practice. The good news? The tongue — and the brain behind it — can be trained.
The Three Pillars of Flavor
When you sip a cup of high-mountain oolong and describe its "flavor," you are unconsciously integrating information from three independent sensory channels. Understanding these channels separately is the first step toward using them deliberately. The neuroscience of flavor perception reveals that what we experience as a single, unified sensation is actually a construction — an elaborate piece of neural engineering assembled in real time from taste, smell, and touch (Hummel & Welge-Lüssen, 2012).
The first pillar is gustation — taste proper — which operates through receptor cells clustered in taste buds on the tongue and palate. These receptors detect only five basic qualities: sweet, bitter, sour, salty, and umami. That's it. Five channels. If you have ever wondered why pinching your nose makes food taste "flat," you have already intuited that taste alone is remarkably limited.
The second pillar is retronasal olfaction — the aroma that reaches your olfactory epithelium not through your nostrils (that is orthonasal olfaction, the sniff before the sip) but from inside your mouth, rising up through the nasopharynx as you chew, swish, and swallow. This single pathway accounts for the vast majority of what we call flavor. Remarkably, the brain processes retronasal and orthonasal signals differently: retronasal odors are routed through the gustatory cortex alongside taste signals, binding them into a unified percept, while orthonasal odors are not (Blankenship et al., 2019). Your brain literally treats aroma-from-the-mouth as part of taste.
The third pillar is the trigeminal nerve system, which detects mechanical and chemical irritation throughout the mouth and nasal cavity. This is not taste and not smell — it is the system responsible for the pucker of astringency, the cooling sensation of menthol, the body and weight of a full liquor, and the tingling warmth of certain spices. In tea, it is arguably the most distinctive pillar of all.
Taste: Five Channels, Infinite Teas
Let us begin on the tongue. Each of the five basic tastes corresponds to a distinct class of receptor proteins, and each plays a specific role in the tea experience.
Sweet and Umami: The T1R Family
Sweetness in tea — the lingering huigan (returning sweetness) prized in Chinese tea evaluation — is detected by the T1R2+T1R3 heteromeric receptor. Umami, the savory, brothy depth that defines shade-grown Japanese teas, is detected by a closely related pair: T1R1+T1R3. These receptors share a subunit (T1R3), which is why sweet and umami can interact and modulate each other (Zhao et al., 2003). The discovery of these receptor families was a watershed moment: it proved that taste qualities are not emergent properties of neural patterns but are encoded at the very first molecular contact between food and body.
For tea lovers, the umami receptor holds particular significance. L-theanine, the amino acid unique to Camellia sinensis and most abundant in shade-grown leaves, directly activates the T1R1+T1R3 receptor. Narukawa et al. (2014) demonstrated this using cell-based assays and site-directed mutagenesis, showing that L-theanine binds to the Venus flytrap domain of T1R1 — the same binding pocket used by glutamate, the prototypical umami compound. This is why a well-made gyokuro or competition-grade matcha can taste uncannily like dashi broth: the same receptor is firing.
Bitter: The Sentinel Sense
Bitterness in tea comes primarily from caffeine and certain catechins. Humans express approximately 25 different bitter taste receptors (the TAS2R family), each tuned to different molecular structures. This is evolution's broadest alarm system — bitterness signals "potentially toxic, proceed with caution." But as any tea drinker knows, bitterness is not simply negative. Balanced bitterness provides structure, complexity, and the contrast that makes sweetness more vivid. In fact, the interplay between the bitterness of catechins and the umami of L-theanine is one of the defining aesthetic tensions in tea (Eldeghaidy et al., 2021).
Sour and Salty
Sourness (acidity) and saltiness play supporting roles in tea. Slight acidity brightens the cup — you notice it in Darjeeling's muscatel character or the lively tang of a sun-dried white tea. Mineral salts from soil contribute subtle salty notes that experienced tasters describe as part of a tea's "terroir signature." Neither dominates, but both shape the background against which other tastes emerge.
Retronasal Olfaction: Where Most of "Flavor" Lives
Here is a claim that surprises most people: if taste provides the skeleton of flavor, retronasal olfaction provides the flesh, the skin, and the clothing. Researchers estimate that 70–80% of what we perceive as flavor is actually aroma detected retronasally. The olfactory epithelium contains roughly 400 types of odorant receptors (compared to taste's five modalities), giving it an enormous palette for discrimination.
When you take a sip of tea, warming the liquid in your mouth causes volatile compounds to vaporize. As you swallow or exhale, these vapors are pushed upward through the nasopharynx to the olfactory epithelium. The brain receives this signal simultaneously with taste input and fuses them into a single experience. Green and colleagues (2012) demonstrated that congruent taste-aroma pairings enhance each other: a sweet taste makes a vanilla retronasal aroma smell sweeter, and the aroma in turn makes the taste seem more intense. These multisensory "flavor objects" are greater than the sum of their parts.
This has direct practical implications for tea tasting. The hundreds of volatile compounds generated during processing — linalool from withering, geraniol from oxidation, pyrazines from roasting — are detected almost entirely through retronasal olfaction. When a professional taster slurps tea loudly, they are not being rude; they are aerating the liquid to maximize volatile release across the palate. When they exhale slowly through the nose after swallowing, they are extending the retronasal window, allowing late-emerging aromatics to register.
"The act of slurping forces air across the surface of the tea in the mouth, volatilizing aroma compounds and driving them into the retronasal passage. It is physics in service of perception."
Sensory science aphorism
Understanding the orthonasal-retronasal distinction also explains a common experience: a tea that smells intensely floral when you inhale the steam from your cup (orthonasal) may taste less floral than expected when you drink it (retronasal). The two routes activate different neural circuits (Blankenship et al., 2019) and may emphasize different components of a complex aroma blend. This is why professional tasters evaluate both the "lid aroma" and the "in-mouth flavor" as separate attributes.
The Trigeminal Dimension: Astringency, Body, and Texture
The third sensory pillar is often the least discussed but the most physically felt. The trigeminal nerve — the largest cranial nerve — threads through the entire oral and nasal cavity, detecting pressure, temperature, and chemical irritation. In tea, its signature sensation is astringency: that dry, puckering, gripping feeling that is emphatically not a taste.
For years, astringency was loosely classified as a tactile sensation caused by tannins precipitating salivary proteins, reducing lubrication in the mouth. That mechanical account is not wrong, but it is incomplete. Schöbel et al. (2014) provided the crucial evidence: using nerve block studies in human subjects, they demonstrated that astringency perception was abolished when the trigeminal nerve was anesthetized but persisted when taste nerves were blocked. Furthermore, they showed that tea catechins — particularly galloylated catechins like EGCG — directly activate trigeminal ganglion neurons via G-protein coupled signaling. Astringency is an active neural signal, not merely a loss of lubrication.
Not all astringency is created equal. Liu et al. (2022) established a hierarchy of astringent potency among tea catechins: ECG > EGCG > GCG > CG > EGC > EC > GC > C. Galloylated catechins (those bearing a gallate ester group) are consistently more astringent than their non-galloylated counterparts. Even more intriguingly, different polyphenols create qualitatively different astringent sensations — catechins produce a "rough" or "grainy" astringency, while flavonol glycosides create a "velvety" or "silky" mouthfeel. This explains why two teas with similar total polyphenol content can feel completely different in the mouth: the ratio of catechin types matters enormously.
Beyond astringency, the trigeminal nerve contributes to the sensation of body — the weight or viscosity of tea in the mouth — and to cooling or warming sensations from certain volatile compounds. The "thick" mouthfeel of a dense aged pu-erh, the "thin" clarity of a high-elevation white tea, the cooling "menthol" finish of certain Wuyi yancha — all are mediated by this third sensory channel.
Your Genes, Your Cup: Variation in Taste Perception
Understanding the three pillars of flavor raises a discomforting question: if two people possess the same receptor systems, why do they describe the same tea so differently? Part of the answer is genetic.
The most studied example is the TAS2R38 gene, which encodes a bitter taste receptor sensitive to compounds containing a thiourea (N–C=S) group. This gene has two common haplotypes: PAV (associated with high bitter sensitivity) and AVI (associated with low sensitivity). Individuals who are PAV/PAV homozygotes — approximately 25% of many populations — are often called supertasters. They experience certain bitter compounds with intense, sometimes overwhelming force. AVI/AVI homozygotes — roughly 25% — are nontasters of these specific compounds. The remaining 50% are heterozygotes with intermediate sensitivity (Campa et al., 2019).
Risso et al. (2016) analyzed TAS2R38 variation across 5,589 individuals from 105 global populations and found evidence of ancient balancing selection — evolution has actively maintained both sensitivity and insensitivity variants in human populations for hundreds of thousands of years, suggesting both confer advantages in different ecological contexts. The global distribution of these haplotypes is not random: population-level differences in bitter sensitivity are real, measurable, and historically shaped.
What does this mean for tea? A supertaster encountering a heavily catechin-rich tea — a rough-processed Assam, an overbrewed sencha — may find the bitterness genuinely unbearable, while a nontaster sipping the same cup perceives only mild bitterness and may instead notice sweetness and body more prominently. Neither is "wrong." They are receiving different neural signals from the same molecules. This has implications for tea preference, brewing habits, and even the cultural evolution of tea preparation methods.
Beyond Genes: Culture, Context, and the Cup
Genetics sets the range; experience tunes the instrument. Cultural background profoundly shapes taste perception — not just preferences, but actual sensory thresholds. Trachootham et al. (2018) demonstrated this vividly in a cross-cultural study comparing Thai and Japanese adults: the Thai group had significantly higher thresholds (lower sensitivity) for all five basic tastes, likely shaped by lifelong exposure to intensely spiced, strongly flavored cuisine. The Japanese group showed markedly greater umami sensitivity — unsurprising in a culture that has cultivated umami-rich ingredients for centuries.
Context matters too. The color of the cup influences perceived flavor intensity. Expectation shapes perception: being told a tea is "rare" or "award-winning" genuinely alters the sensory experience by priming attention to positive attributes. Mood, fatigue, hydration, and the time of day all modulate receptor sensitivity. Even the altitude at which you are tasting — by affecting olfactory receptor function — can shift flavor perception.
None of this makes sensory evaluation unreliable. It makes it human. The solution is not to pretend these variations do not exist but to develop a shared vocabulary and structured method that allows people with different sensory equipment to communicate meaningfully about what they experience.
Building a Tasting Vocabulary
Professional tea evaluation traditions around the world converge on a common insight: subjective experience becomes communicable when structured by systematic vocabulary. The goal is not to replace personal experience with jargon but to give you tools for noticing more, remembering more, and sharing more precisely.
The Five Dimensions of Evaluation
Across Chinese gongfu evaluation, British trade cupping, and Japanese tea ceremony assessment, five dimensions appear consistently:
- Appearance — the color, clarity, and viscosity of the liquor; the shape and color of dry and wet leaves.
- Aroma — both orthonasal (lid aroma, steam aroma) and retronasal (in-mouth fragrance). Described using analogy: floral, vegetal, roasted, fruity, earthy, marine, mineral.
- Taste — the five basic tastes, their intensity, and their sequence over time.
- Mouthfeel — trigeminal sensations: astringency (rough, silky, velvety), body (thin, medium, thick), and textural qualities (oily, crisp, chalky).
- Finish — the aftertaste and how it evolves. Chinese tasters prize huigan (returning sweetness), chaqi (the bodily sensation of the tea), and the duration of flavor after swallowing.
Zhu et al. (2019) developed a 27-term lexicon for Hunan fuzhuan brick tea using trained panelists and quantitative descriptive analysis, demonstrating that even complex, fermented dark teas can be consistently evaluated when assessors share a calibrated vocabulary. Their work showed that both traditional Chinese cupping methods and modern sensory science approaches achieved high inter-rater consistency when vocabulary was standardized.
Tradition-Specific Approaches
Chinese evaluation emphasizes the temporal arc of the experience — how the tea changes from first infusion to fifth, and from the initial sip through the lingering aftertaste. The concepts of huigan, yanyun (rock rhyme, in Wuyi oolong), and chaqi (tea energy) extend evaluation beyond the purely sensory into the bodily and even the aesthetic.
British trade cupping prioritizes consistency, defect detection, and grading. Leaves are weighed precisely, water is standardized, steeping time is exact. The vocabulary focuses on manufacturing quality: "brisk" (a positive astringency indicating fresh oxidation), "bakey" (overheated during firing), "malty" (a desirable character in Assam).
Japanese evaluation places umami at the center. In competition gyokuro judging, a rich, savory, almost marine depth is the highest quality marker — a sensory ideal that directly reflects the L-theanine content built up through shade growing, which, as we have seen, activates the T1R1+T1R3 umami receptor (Narukawa et al., 2014).

From Knowledge to Practice
Sensory literacy is a skill, not a talent. Research consistently shows that trained tasters do not have "better" taste buds — they have better attention and better vocabulary. Training reshapes the cognitive frameworks you apply to sensory data, not the data itself. A novice and a master tasting the same aged sheng pu-erh receive the same neural signals; the master has learned to parse those signals into meaningful categories, to notice the camphor note beneath the earthiness, to track the astringency as it softens across steepings.
This is good news. It means your sensory world can expand. Every time you taste tea deliberately — pausing to separate aroma from taste from mouthfeel, naming what you notice, comparing your notes with others — you are building the neural architecture of expertise. The compounds are already in the cup. The receptors are already on your tongue. What we are training is the bridge between sensation and understanding.
Key Takeaways
- "Flavor" is not a single sense but a construction from three distinct systems: gustation (five basic tastes), retronasal olfaction (hundreds of volatile compounds), and trigeminal sensation (astringency, body, texture).
- Retronasal olfaction — aroma rising from inside the mouth — contributes the majority of perceived flavor and is processed by the gustatory cortex, unlike orthonasal smell.
- Specific tea compounds map to specific receptors: L-theanine activates T1R1+T1R3 (umami), catechins activate trigeminal neurons (astringency), and volatile compounds reach olfactory receptors retronasally.
- Genetic variation in TAS2R38 creates genuine differences in bitter perception — supertasters and nontasters experience the same tea differently at the receptor level.
- Cultural exposure, context, and expectation further modulate perception across all five basic tastes, as demonstrated in cross-cultural research.
- A structured evaluation framework — appearance, aroma, taste, mouthfeel, finish — enables meaningful communication about subjective sensory experience.
- Sensory expertise is built through deliberate attention and vocabulary development, not through innately "better" taste buds.
Now that you can name what you taste and understand why others may taste differently, we turn in Chapter 5 to the cultural architectures built around tea tasting. From the meditative silence of Japanese chanoyu to the raucous communality of Moroccan mint tea service, every tea culture has developed rituals that frame the sensory experience you've just learned to parse. The question shifts from "What am I tasting?" to "What does this tasting mean?"
References
Blankenship, M. L., Grigorova, M., Katz, D. B., & Maier, J. X. (2019). Retronasal odor perception requires taste cortex, but orthonasal does not. Current Biology, 29(1), 62–69.e3.
Campa, D., Mottini, A., Sereno, M., et al. (2019). TAS2R38 bitter taste receptor and attainment of exceptional longevity. Scientific Reports, 9, 18047.
Eldeghaidy, S., Marciani, L., Hort, J., et al. (2021). Mechanisms of umami taste perception: From molecular level to brain imaging. Critical Reviews in Food Science and Nutrition, 63(4), 525–540.
Green, B. G., Nachtigal, D., Hammond, S., & Lim, J. (2012). Enhancement of retronasal odors by taste. Chemical Senses, 37(5), 469–479.
Hummel, T., & Welge-Lüssen, A. (2012). Retronasal perception of odors. Physiology & Behavior, 107(4), 484–487.
Liu, Z., de Bruijn, W. J. C., Bruins, M. E., & Vincken, J.-P. (2022). Exploring the relative astringency of tea catechins and distinct astringent sensation via an in vitro assay. Foods, 11(18), 2807.
Narukawa, M., Toda, Y., Nakagita, T., Hayashi, Y., & Misaka, T. (2014). L-Theanine elicits umami taste via the T1R1 + T1R3 umami taste receptor. Amino Acids, 46(6), 1583–1587.
Risso, D. S., Mezzavilla, M., Pagani, L., Robino, A., Morini, G., et al. (2016). Global diversity in the TAS2R38 bitter taste receptor. Scientific Reports, 6, 25506.
Schöbel, N., Radtke, D., Kyereme, J., et al. (2014). Astringency is a trigeminal sensation that involves the activation of G protein–coupled signaling by phenolic compounds. Chemical Senses, 39(6), 471–487.
Trachootham, D., Satoh-Kuriwada, S., Lam-ubol, A., et al. (2018). Differences in taste perception and spicy preference: A Thai–Japanese cross-cultural study. Chemical Senses, 43(1), 65–74.
Zhao, G. Q., Zhang, Y., Hoon, M. A., et al. (2003). The receptors for mammalian sweet and umami taste. Cell, 115(3), 255–266.
Zhu, Y., Dong, L., Huang, L., et al. (2019). Lexicon development and quantitative descriptive analysis of Hunan fuzhuan brick tea infusion. Food Research International, 120, 292–301.