The Architecture of a Night
What Happens After You Close Your Eyes
Sleep is not a single state. It is a structured sequence of biologically distinct stages, and the balance between them shifts dramatically across the night — cut from one end, and you lose different capacities than if you cut from the other.
Imagine two students, Maya and Jake, both getting exactly six hours of sleep before an exam. Maya goes to bed at midnight and sets her alarm for 6 a.m. Jake stays up until 3 a.m. and sleeps until 9 a.m. They sleep for the same duration — but Maya wakes up feeling foggy and emotionally raw, while Jake feels physically sluggish but mentally sharp. Same total hours, radically different outcomes. Why?
The answer lies in a fact that surprises most people: sleep is not one thing. It is a structured sequence of biologically distinct stages, each performing different work, and the balance between those stages shifts dramatically across the night. Cut from one end, and you lose different capacities than if you cut from the other. Understanding this architecture — this hidden blueprint of a night — changes how you think about every sleep decision you make.
Sleep Is Not a Single State
When you close your eyes and drift off, your brain does not simply power down like a laptop entering standby mode. Instead, it begins executing a complex, highly organised sequence of neural states that cycle roughly every 90 minutes throughout the night. This recurring pattern is called the ultradian rhythm — a within-sleep rhythm nested inside the 24-hour circadian rhythm you studied in the previous lesson.
Sleep scientists divide this sequence into two broad categories: non-rapid eye movement sleep, which itself has three stages called N1, N2, and N3, and rapid eye movement sleep, the stage most associated with vivid dreaming. A single pass through all stages constitutes one sleep cycle, and a typical eight-hour night contains four to six of these cycles (Patel et al., 2024). But here is the critical insight that will frame this entire chapter: the cycles are not identical copies. Early cycles are dominated by deep NREM sleep; later cycles are dominated by REM. This asymmetry has profound consequences.
Descending the Staircase — NREM Sleep
N1 — the hypnagogic threshold
The lightest stage of sleep, N1, is the brief transition between wakefulness and sleep — typically lasting only one to five minutes. Your brain shifts from the alpha waves of relaxed wakefulness to the slower theta waves of early sleep. Muscles begin to relax, and you may experience hypnagogic phenomena: fleeting sensory hallucinations, the sensation of falling, or sudden whole-body muscle jerks called hypnic myoclonia (Patel et al., 2024).
N1 is so light that most people, if awakened from it, will deny having been asleep at all. It constitutes only about 5% of total sleep time and is not considered restorative in itself. Think of it as a doorway — not a room you spend much time in, but one you must pass through repeatedly as you transition between deeper stages across the night.
N2 — the workhorse of sleep
You spend more time in N2 than any other stage — roughly 45–55% of total sleep time. And while it has historically been treated as a mere waypoint between light and deep sleep, recent research has revealed N2 to be a remarkably active processing stage in its own right, thanks to two distinctive electrical signatures: sleep spindles and K-complexes.
Sleep spindles are brief bursts of rhythmic neural activity (11–16 Hz) generated by the thalamus. They appear as tight, waxing-and-waning oscillations on an EEG readout, and they are far more than electrical noise. In a landmark study, Nishida and Walker (2007) demonstrated that improvements in motor skill memory — the kind of memory involved in learning a piano sequence or a tennis serve — correlated specifically with the density of sleep spindles during N2 sleep. More strikingly, these spindles were regionally specific: if a participant learned a right-hand motor task, spindle density increased over the left motor cortex — the exact hemisphere controlling that hand. The brain was not just passively resting; it was rehearsing the day's motor learning at an anatomically precise location.
K-complexes, the other hallmark of N2, are large, sharp waveforms that appear to serve a gating function. When an external sound occurs during N2 sleep — a door closing, a car horn — a K-complex fires in response, essentially evaluating whether the stimulus warrants waking up. If the sound is deemed non-threatening, sleep continues. K-complexes thus function as a night-watch system, filtering irrelevant noise while protecting the continuity of sleep (Patel et al., 2024).
N3 — deep sleep and the rebuilding crew
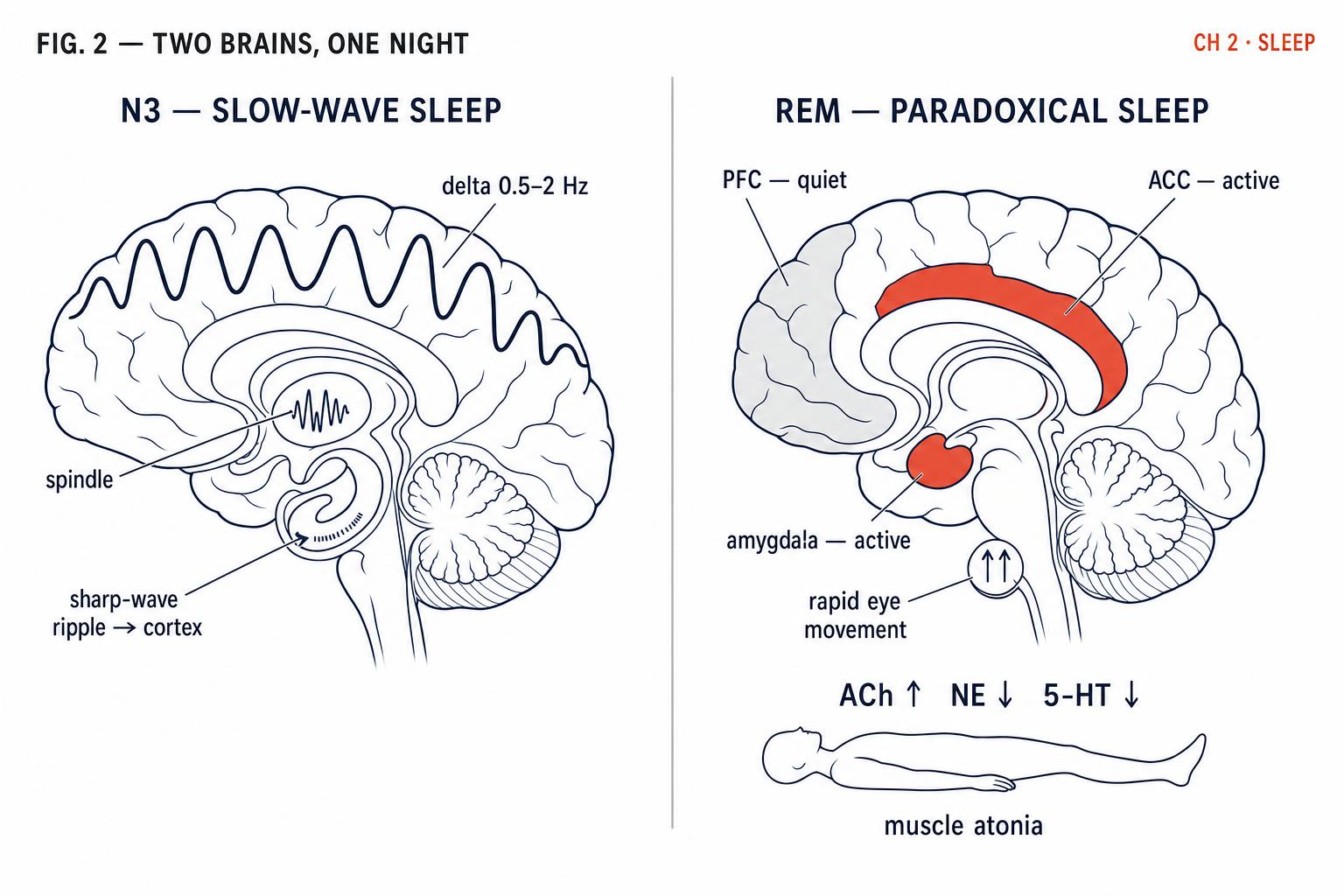
N3, also called slow-wave sleep, is the deepest stage of NREM sleep — and, in many ways, the most physiologically powerful. It is characterised by high-amplitude, low-frequency delta waves (0.5–2 Hz) that sweep across the cortex in slow, synchronised oscillations. This is the stage from which you are hardest to wake, and the stage in which your brain and body perform some of their most critical maintenance work.
Physiologically, N3 is when the body releases the majority of its daily growth hormone, driving tissue repair, muscle recovery, and immune system strengthening (Patel et al., 2024). This is why athletes who lose deep sleep see measurable declines in physical recovery — a connection we will explore further in a later chapter.
But N3's contributions extend far beyond the body. The active system consolidation model, proposed by Diekelmann and Born (2010), describes a remarkable neural process that unfolds during slow-wave sleep. Here is how it works: memories that were initially encoded in the hippocampus during the day are spontaneously reactivated during slow-wave sleep. These reactivations occur during sharp wave-ripples in the hippocampus, which are temporally coordinated with thalamic sleep spindles and the slow cortical oscillations that define N3. This triple-nested coordination — ripple within spindle within slow oscillation — creates a neural conveyor belt that gradually transfers memory representations from the hippocampus to more stable, long-term storage sites in the neocortex (Rasch & Born, 2013).
This process particularly benefits declarative memories — memories for facts, events, and explicit knowledge. In experimental paradigms, participants who studied word pairs or spatial layouts before sleep showed significantly better recall after sleep rich in slow-wave activity, compared to equivalent periods of wakefulness (Diekelmann et al., 2009). The effect was especially strong for weakly encoded items — precisely the kind of fragile new knowledge that needs consolidation most.
The sleeping brain is not simply maintaining memories — it is actively reorganising them, extracting their gist, and integrating them into existing knowledge networks.
Diekelmann & Born (2010)
More recently, researchers have linked N3 to the glymphatic system — a waste-clearance mechanism in which cerebrospinal fluid flushes through brain tissue, clearing metabolic byproducts including amyloid-beta, a protein implicated in Alzheimer's disease. While this research is still emerging and the precise stage-specificity is under active investigation, the current evidence suggests that deep NREM sleep provides the optimal conditions for this neural housekeeping.
The Paradox of REM Sleep
After roughly 70–90 minutes of progressively deepening NREM sleep, something extraordinary happens. The brain reverses course and enters rapid eye movement sleep — a stage so physiologically distinct from everything that came before that its discoverers initially called it "paradoxical sleep." The paradox: your brain becomes almost as electrically active as when you are awake, yet your body becomes almost completely paralysed.
During REM, the brainstem sends inhibitory signals to motor neurons throughout the body, producing a state called muscle atonia. Your voluntary muscles are effectively disconnected. This paralysis appears to be a protective mechanism — preventing you from physically acting out the vivid, story-like dreams that characterise this stage. When the atonia mechanism fails, the result is REM sleep behaviour disorder, in which people kick, punch, or leap from bed during dreams.
Meanwhile, the brain is extraordinarily busy. The emotional centres — particularly the amygdala and the anterior cingulate cortex — become highly active, while the prefrontal cortex, responsible for logical oversight and impulse control, becomes relatively quiet. This unique neurochemical environment — high in acetylcholine, low in norepinephrine and serotonin — appears to create ideal conditions for emotional memory processing.
Goldstein and Walker (2014) demonstrated that sleep deprivation amplified amygdala reactivity to negative emotional stimuli by roughly 60%, accompanied by reduced connectivity between the amygdala and the medial prefrontal cortex — the neural circuit that normally keeps emotional responses in check. Their findings suggest that sleep, and REM sleep in particular, plays a role in recalibrating emotional responses overnight.
A note on nuance — what REM does and doesn't do
It is tempting to assign REM sleep sole responsibility for emotional regulation — and some popular accounts do exactly that, claiming that without sufficient REM sleep, you become essentially emotionally irrational. The evidence, however, is more nuanced than this narrative suggests.
Tempesta et al. (2018) conducted a comprehensive review of sleep and emotional processing, concluding that while evidence converges on REM sleep benefiting emotional processing, "contradictory findings exist and crucial aspects remain unclear." Total sleep deprivation studies consistently show emotional dysregulation, but isolating the specific contribution of REM versus NREM remains methodologically challenging. Some emotional memory benefits appear even after short naps dominated by NREM sleep (Diekelmann et al., 2009).
Schönauer and Rasch (2015) proposed a more integrated model: NREM slow-wave sleep initially consolidates the memory content of emotional experiences, while REM sleep subsequently integrates those consolidated traces into broader neocortical networks, gradually decontextualising them — separating the emotional charge from the factual content. This two-stage model is elegant, but it also makes clear that no single stage works alone. The lesson for students of sleep science is important: be cautious of any claim that assigns a complex psychological function to a single sleep stage. Sleep stages work as a coordinated system.
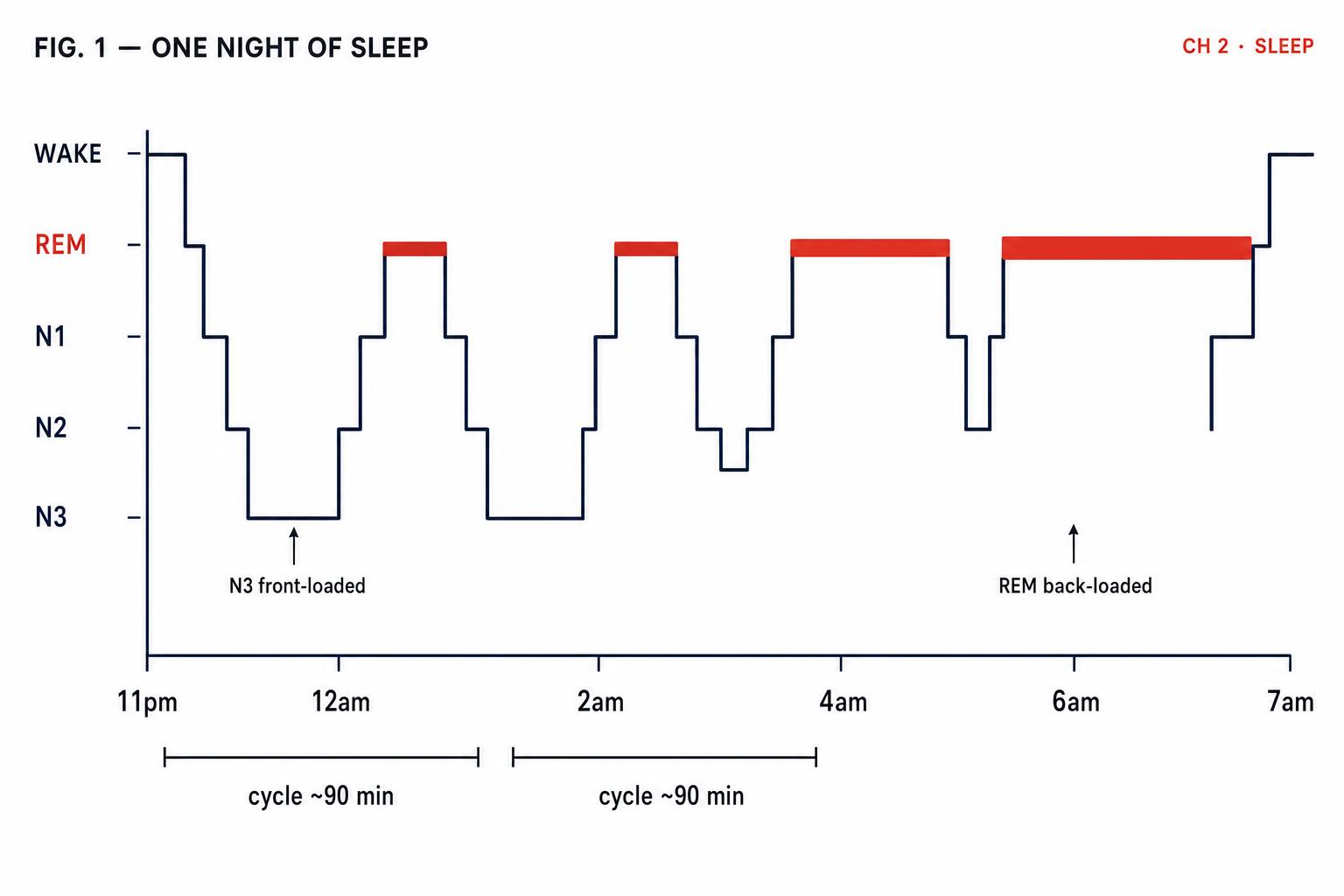
The Shifting Architecture of a Night
Now that you understand what each stage does individually, we can examine how they combine across a full night — and why the sequence matters as much as the stages themselves.
A typical night of eight hours contains four to six complete sleep cycles, each lasting approximately 90 minutes. But these cycles are not interchangeable. The first two cycles — roughly the first three hours of sleep — are dominated by deep N3 slow-wave sleep, with only brief periods of REM. As the night progresses, N3 diminishes and REM periods grow progressively longer. By the final cycle — typically between 6 and 8 a.m. for someone who fell asleep around 11 p.m. — REM may last 40–60 minutes, while N3 has nearly vanished (Patel et al., 2024).
This asymmetry is not random. It is driven by the two regulatory processes you learned about in the previous lesson. Process S, homeostatic sleep pressure — primarily adenosine accumulation — is at its peak when you first fall asleep, and it is this intense pressure that drives the brain down into deep N3 sleep early in the night. As N3 sleep dissipates adenosine, Process S weakens. Meanwhile, Process C, the circadian signal, increasingly favours REM sleep as the night progresses, with REM propensity peaking in the early morning hours. The two processes together sculpt the shifting landscape of your sleep cycles.
This brings us back to Maya and Jake from our opening scenario. Maya, sleeping from midnight to 6 a.m., captured most of her deep N3 sleep — front-loaded in the first three hours — but lost her final, REM-rich cycles. Jake, sleeping from 3 a.m. to 9 a.m., slept through the circadian REM window but missed the initial deep sleep surge — though his high sleep pressure would have partially compensated by driving rapid descent into N3. The result: Maya preserved physical restoration but sacrificed emotional processing; Jake got more REM but less total deep sleep. Neither got a complete night's architecture.
Why Architecture Matters — Practical Implications
Understanding sleep architecture transforms abstract neuroscience into practical decision-making. Here are three implications that will recur throughout this course.
Caffeine timing reshapes your stages. Caffeine blocks adenosine receptors — the very mechanism that drives Process S and the descent into deep N3 sleep. Consuming caffeine in the afternoon or evening does not merely delay sleep onset; it selectively reduces deep slow-wave sleep even when total sleep duration is preserved. You will explore this mechanism in detail in a later chapter, but the architectural logic should already be clear: anything that blunts adenosine's signal will disproportionately erode the N3-rich early cycles.
Nap design depends on which stage you need. A 20-minute nap captures mostly N2, gaining you spindle-mediated motor consolidation and mild restoration. A 90-minute nap allows a complete cycle including N3 and possibly REM, offering deeper restoration and broader memory benefits — but also risks sleep inertia upon waking. When you study nap architecture in a later chapter, you will design naps based on which stage's functions you most need at that moment.
Exercise timing interacts with deep sleep. Vigorous exercise increases N3 sleep — but only when performed with appropriate timing relative to circadian phase. Evening exercise too close to bedtime can elevate core body temperature and delay sleep onset, paradoxically reducing the N3 it would otherwise enhance. A later chapter will explore the optimal timing window.
Connecting the Models — Process S, Process C, and Sleep Stages
In the previous lesson, you learned that Process S (homeostatic sleep pressure) and Process C (circadian alerting signal) jointly determine when you sleep. Now you can see that they also determine how you sleep. Process S, driven by adenosine accumulation, powers the deep slow-wave sleep that dominates early cycles. As N3 discharges that pressure, Process C's circadian gate swings open for REM. The two processes are not just scheduling sleep — they are programming its internal structure.
This interconnection means that anything disrupting Process S or Process C does not just change sleep timing; it changes sleep composition. Shift workers, jet-lagged travellers, and students pulling all-nighters do not merely sleep fewer hours — they sleep differently configured hours, with potentially altered ratios of N3 to REM. The architectural lens makes visible what a simple "hours slept" metric cannot.
Key Takeaways
- Sleep is not a uniform state but a structured sequence of distinct stages — N1, N2, N3, and REM — cycling roughly every 90 minutes in an ultradian rhythm.
- N2 sleep spindles consolidate motor memories at regionally specific brain locations, while K-complexes filter external stimuli to protect sleep continuity.
- N3 (slow-wave sleep) is critical for growth hormone release, immune function, glymphatic waste clearance, and the active consolidation of declarative memories through coordinated hippocampal replay.
- REM sleep is associated with emotional memory processing and affective recalibration, but its specific contribution is more nuanced than popular claims suggest — NREM and REM stages work as a coordinated system.
- Sleep architecture shifts across the night: N3 dominates the first half, REM dominates the second. Truncating sleep from different ends disproportionately costs different functions.
- Process S drives early deep sleep; Process C gates later REM sleep — the two-process model from Chapter 1 directly programs the internal structure of each night.
- Practical decisions about caffeine, nap timing, exercise, and sleep schedules all reshape sleep architecture by selectively affecting specific stages.
You now know the SCN sets the clock and that each night is a sculpted architecture of stages. Next chapter: the single most powerful tool for controlling that clock — light. We'll meet the melanopsin-containing retinal cells that hand light directly to the SCN, the phase-response curve that decides whether a given dose of light pushes you earlier or later, and the practical protocols (morning sunlight; evening avoidance) that follow. Light, treated correctly, is the most powerful button on your clock.
References
Diekelmann, S., & Born, J. (2010). The memory function of sleep. Nature Reviews Neuroscience, 11(2), 114–126.
Diekelmann, S., Wilhelm, I., & Born, J. (2009). The whats and whens of sleep-dependent memory consolidation. Sleep Medicine Reviews, 13(5), 309–321.
Goldstein, A. N., & Walker, M. P. (2014). The role of sleep in emotional brain function. Annual Review of Clinical Psychology, 10, 679–708.
Nishida, M., & Walker, M. P. (2007). Daytime naps, motor memory consolidation and regionally specific sleep spindles. PLoS ONE, 2(4), e341.
Patel, A. K., Reddy, V., Shumway, K. R., & Araujo, J. F. (2024). Physiology, sleep stages. In StatPearls. StatPearls Publishing.
Rasch, B., & Born, J. (2013). About sleep's role in memory. Physiological Reviews, 93(2), 681–766.
Schönauer, M., & Rasch, B. (2015). The role of REM sleep theta activity in emotional memory. Frontiers in Psychology, 6, 1439.
Tempesta, D., Socci, V., De Gennaro, L., & Ferrara, M. (2018). Sleep and emotional processing. Sleep Medicine Reviews, 40, 183–195.