Understanding Your Sleep: The Science of Rest, Rhythm, and Recovery
This course tells a single, unfolding story: your body runs on a clock, that clock shapes the architecture of every night, and a handful of powerful levers — light, caffeine, movement, stimulation, and strategic rest — c…
Your Internal Clock — The 24-Hour Rhythm You Never Chose
Every student walks into this course already running a biological clock they've never consciously set. This opening chapter reveals that clock — the suprachiasmatic nucleus (SCN), a tiny cluster of roughly 20,000…
Should 8am classes be illegal? Like, actually against the law?
Controversial take
Would you rather: Take all your classes starting at 7am OR all your classes starting at 7pm?
Would you rather
Hot take: Being a 'morning person' isn't a personality trait — you just won the schedule lottery.
Hot take
Is making night owls show up at 8am basically discrimination? Discuss.
Real talk
Can you actually train yourself to function on 5 hours of sleep, or are you just in denial about slowly falling apart?
Asking for a friend
Which is worse: Being forced to wake up 3 hours earlier than your body wants, or staying awake 3 hours later than your body wants?
Quick poll
Chronotype Speed Dating
10-12 minutesStudents turn to neighbors (left, right, front, back as possible) and conduct 2-minute micro-interviews: 'When do you naturally wake without an alarm on free days?' 'When do you feel most alert?' 'When would you schedule an important exam if you could choose?' After 3-4 quick interviews, instructor polls the room by asking larks (morning types) to stand, then neutrals, then owls (evening types). Discuss the visible distribution and what it reveals about chronotype diversity. Follow with: 'Keep standing if you currently have a morning class that conflicts with your natural rhythm.' Visual representation of social jetlag.
Social Jetlag Calculator Pairs
8-10 minutesStudents pair with a neighbor. Provide formula on screen: Social Jetlag = |Midpoint of sleep on free days - Midpoint of sleep on work days|. Example: Sleep 11pm-7am on weekdays (midpoint 3am), but midnight-10am on weekends (midpoint 6am) = 3 hours social jetlag. Pairs calculate their own values and discuss: What causes your jetlag? What would it take to reduce it? Instructor then polls: 'Raise hands if your social jetlag is 0-1 hour... 1-2 hours... 2+ hours.' Discuss health implications of chronic misalignment.
Zeitgeber Power Rankings Debate
15-18 minutesPresent 6 zeitgebers on screen: Light, Meal timing, Exercise, Social interaction, Temperature, Alarm clocks. Students discuss with 2-3 neighbors (turn and talk in seats): 'Rank these from most to least powerful for entraining YOUR circadian rhythm.' After 3-4 minutes, conduct a live poll by rows: 'Row 1-3, who chose light as #1? Rows 4-6, who chose meal timing?' etc. Instructor identifies 2-3 disagreement points. Ask volunteers to defend their rankings (quick 30-second defenses). Reveal evidence hierarchy: Light dominates, but social zeitgebers matter for adolescents. Discuss why alarm clocks are weak zeitgebers (no phase-shifting properties).
Process S vs Process C Thought Experiment
6-8 minutesInstructor narrates a scenario: 'You've been awake for 16 hours. It's 11pm. Your sleep pressure (Process S) is screaming GO TO BED. But then you walk outside into bright moonlight and cool air. What happens to your alertness?' Students discuss with neighbors: Which process wins? Why? Instructor reveals: Circadian alerting (Process C) can temporarily override high sleep pressure. Present second scenario: 'You pulled an all-nighter (Process S maxed out), but it's now 10am and you're in bright light. Why don't you immediately collapse?' Students predict, then discuss the two-process interaction. Use hand-raise poll: 'Who has experienced a second wind late at night? That's Process C temporarily winning.'
SCN Damage Diagnosis Challenge
12-15 minutesPresent 3 patient case studies on screen (fictional but based on research). Case 1: Patient sleeps at random times, sometimes 3am, sometimes 8pm, no pattern. Case 2: Patient always sleeps 2am-10am regardless of work schedule or light exposure. Case 3: Patient's sleep shifts 1 hour later each day (free-running). Students discuss with neighbors: Which suggests SCN damage? Which suggests intact SCN but poor entrainment? Which might be a chronotype issue? After 4-5 minutes, instructor polls the room for each case. Reveal answers: Case 1 = SCN damage (no endogenous rhythm), Case 2 = Delayed chronotype (intact but shifted rhythm), Case 3 = Entrainment failure (functioning oscillator, no zeitgeber input - like blind individuals). Discuss what this reveals about SCN's role as the master clock.
Design Your Ideal Schedule
10-12 minutesStudents sketch a 24-hour wheel with a neighbor, marking their ideal schedule if society had no constraints: wake time, peak productivity hours, meal times, social time, sleep onset. Compare to their ACTUAL schedule. Identify conflicts. Instructor asks: 'Raise your hand if your ideal wake time is 2+ hours different from reality.' Then: 'Keep your hand up if this mismatch causes problems.' Leads to discussion: Why does society impose a narrow schedule? What are the costs of chronotype discrimination? Could we design schools/workplaces to accommodate circadian diversity? Volunteers share one change that would improve their circadian alignment.
Transcript
It's two A-M on a Tuesday, and you're wide awake. Your roommate, who went to bed at ten thirty, is sleeping soundly. You've tried everything — putting the phone away, counting breaths, lying still in the dark — but your body simply refuses to feel tired. Meanwhile, that same roommate will spring out of bed at six fifteen without an alarm, chipper and alert, while you'll drag yourself to your nine A-M lecture feeling like you're moving through wet concrete. Is something wrong with you? Are you lazy? Do you lack discipline?
[short pause]
The answer, as it turns out, has almost nothing to do with willpower. You are running a biological clock — one you never chose, never set, and cannot simply override by wishing it so. In this chapter, we'll open the case of that clock, examine the machinery inside, and discover why understanding it may be one of the most practically useful things you learn in college.
[short pause]
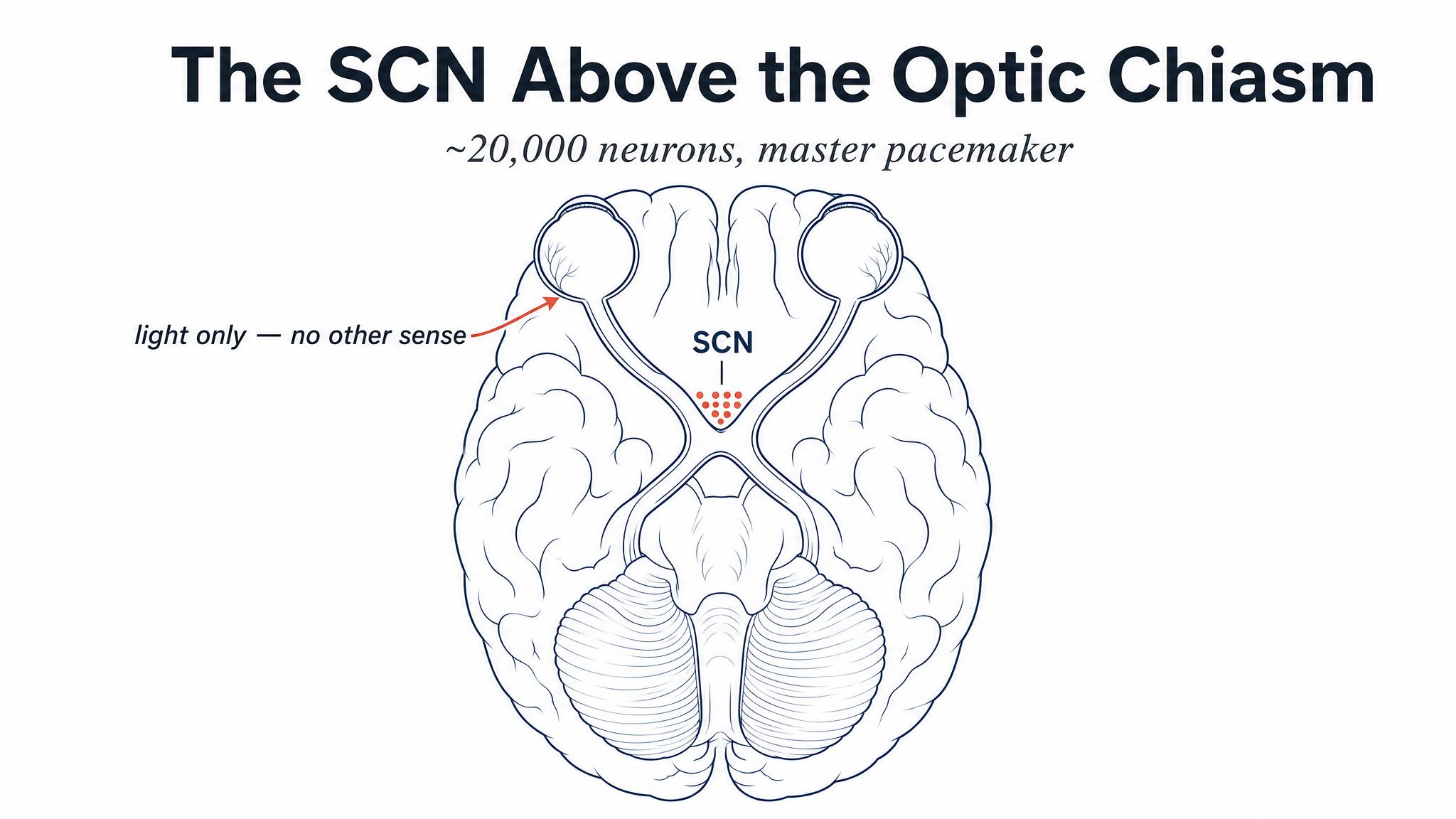
Deep inside your brain, just above the point where your optic nerves cross on their way from your eyes to your visual cortex, sits a structure smaller than a grain of rice. It's called the SUPRACHIASMATIC NUCLEUS, or S-C-N, and it contains roughly twenty thousand neurons — a vanishingly small fraction of the brain's eighty-six billion, yet arguably among the most influential. As Hastings and colleagues reported in 2018, these neurons are your master CIRCADIAN clock, the biological pacemaker that orchestrates a twenty-four-hour rhythm of wakefulness, sleepiness, body temperature, hormone secretion, and dozens of other physiological processes.
The word circadian comes from the Latin circa, meaning "about," and diem, meaning "day." That "about" is critical. In carefully controlled laboratory conditions where people live without clocks, windows, or any sense of external time, the human circadian cycle averages approximately twenty-four point two hours — slightly longer than a full day. According to Hastings and colleagues in 2018, left entirely to its own devices, your internal clock would gradually drift later and later, like a watch that runs a few minutes slow. You'd fall asleep a little later each night, wake a little later each morning, and slowly rotate around the clock over the course of weeks.
This doesn't happen in normal life, of course, because each day your S-C-N receives corrective signals from the environment that reset it to exactly twenty-four hours. Think of it like nudging that slow watch back to the correct time every morning. These environmental corrective signals have a name — a wonderfully evocative German word: ZEITGEBERS, meaning "time givers."
[short pause]
The S-C-N's position above the optic chiasm is not an accident of anatomy — it's the architecture of a system designed around light. A specialized subset of retinal cells called intrinsically photosensitive retinal ganglion cells don't contribute to vision in the way rods and cones do. Instead, they detect the overall brightness and blue-wavelength content of ambient light and send that information directly to the S-C-N via a dedicated neural highway called the retinohypothalamic tract. As Hastings and colleagues found in 2018, this is how your clock knows whether it's day or night — not because you've looked at a clock on the wall, but because photons from the environment are physically reaching these cells and signaling to the S-C-N.
When bright light hits the S-C-N during the biological morning, it advances the clock slightly, correcting for that natural tendency to run slow. When light hits the S-C-N in the biological evening, it can delay it. This daily resetting process is called ENTRAINMENT — the synchronization of an internal oscillator to an external cycle. Light is the most powerful zeitgeber, but it is not the only one.
[short pause]
While light dominates the entrainment hierarchy, your circadian system also responds to several nonphotic zeitgebers — cues that have nothing to do with brightness. Quante and colleagues in 2019 assessed the relative influence of meal timing, physical activity, and light exposure on rest-activity patterns using wrist actigraphy and smartphone tracking. Their findings confirmed what decades of animal research had suggested: the timing of meals, bouts of physical activity, and even social interactions all contribute to synchronizing your biological clock with the external world.
Consider what this means practically. When you eat breakfast at the same time each morning, your digestive system "expects" that meal, pre-releasing enzymes and priming insulin sensitivity. When you exercise at a consistent hour, your cardiovascular system anticipates the demand. When you regularly encounter other people — chatting with a barista, attending a lecture, meeting friends for dinner — those social rhythms reinforce the temporal structure of your day. Each of these is a zeitgeber, each a gentle nudge telling your S-C-N, "It's this time of day."
[short pause]
Before moving forward, consider every cue in your daily life that might tell your body what time it is. Think beyond your alarm clock — what about meals, light exposure, social contact, exercise? Which of these do you think is the most powerful signal, and why? You may be surprised by which cues your circadian system cares about most — and which ones it largely ignores.
[short pause]
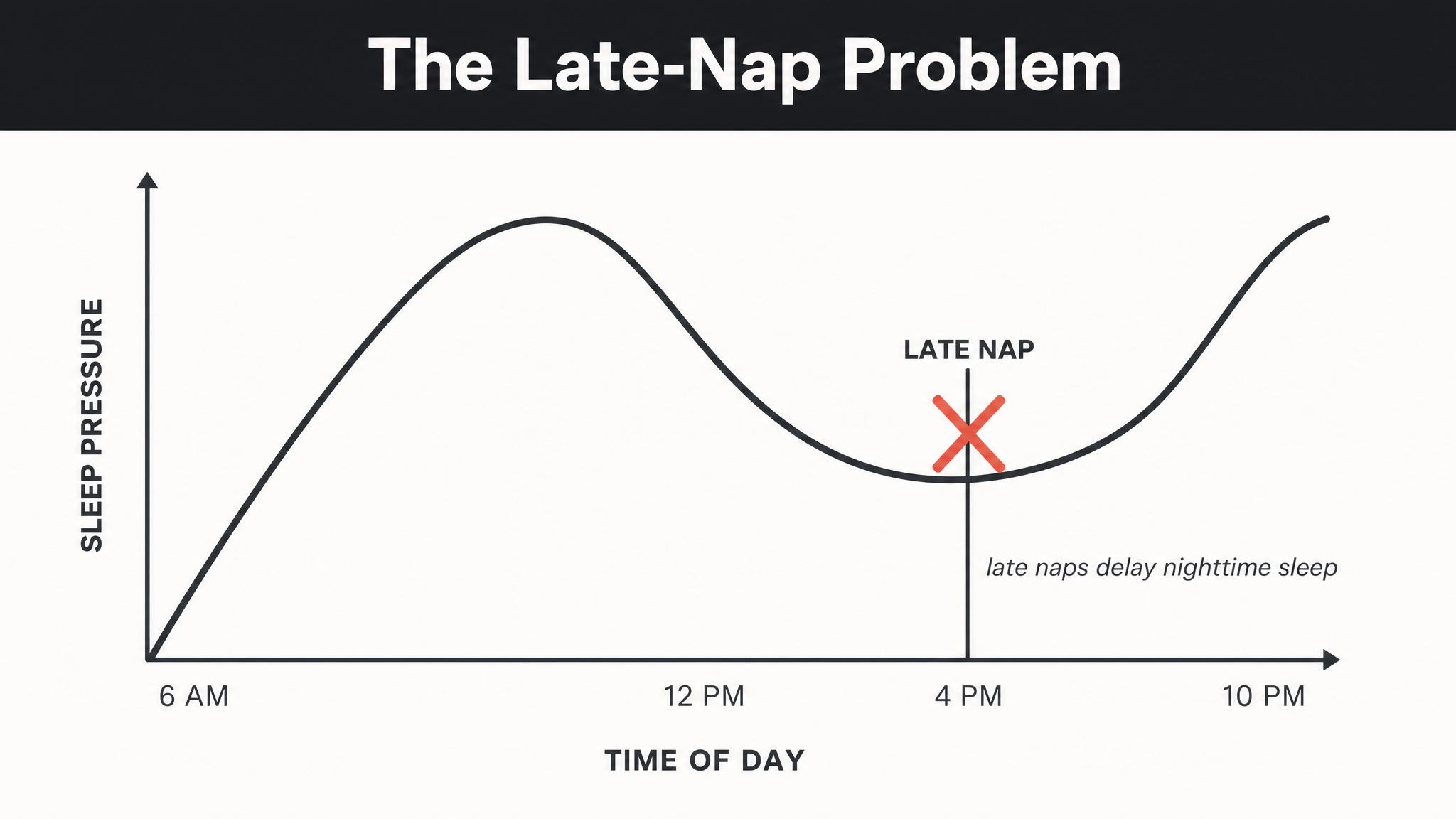
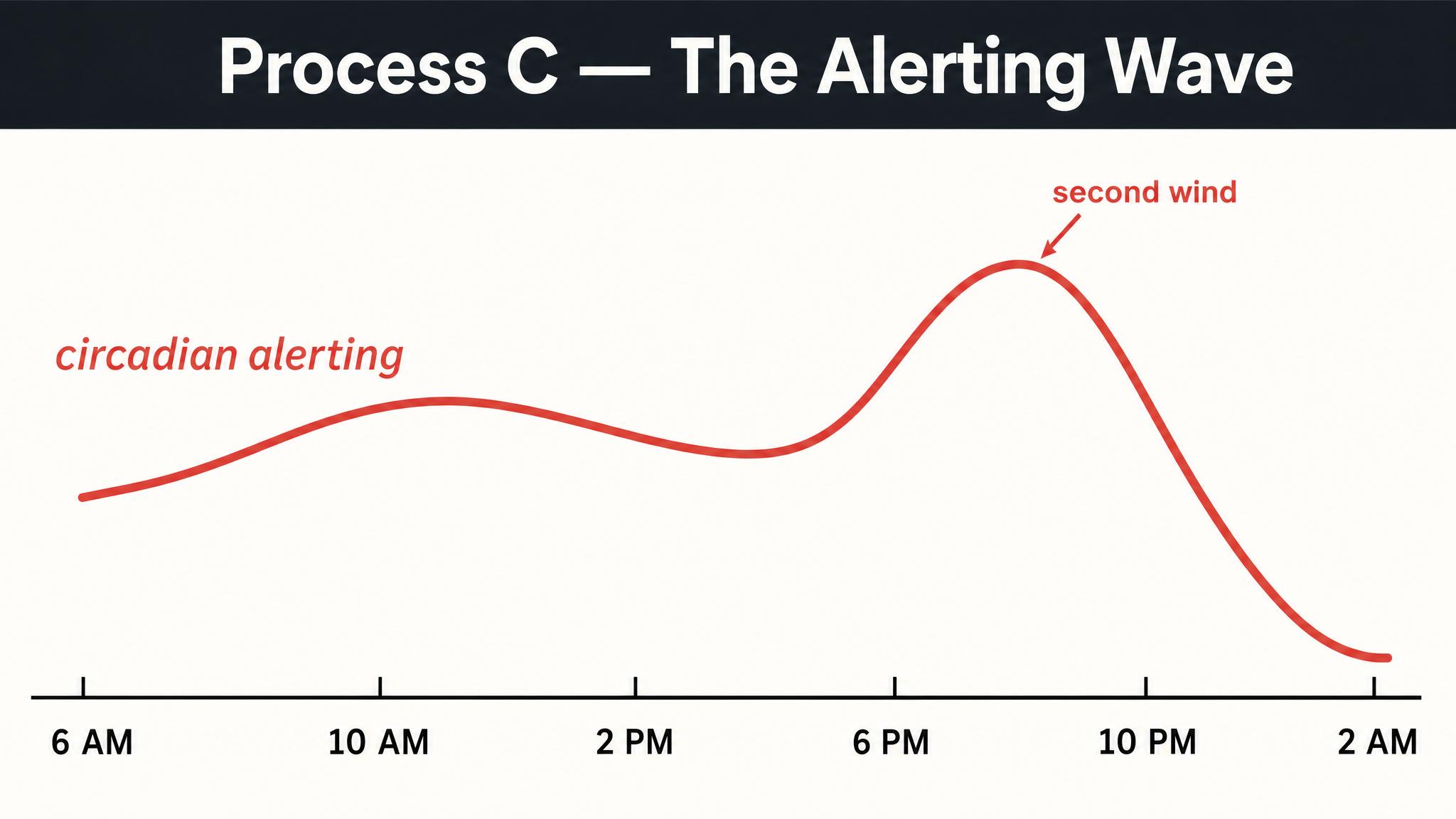
Understanding that you have a circadian clock is a good start, but it doesn't fully explain when you feel sleepy. After all, you've probably experienced being exhausted at three P-M but catching a second wind by six P-M — even though you didn't nap. If sleepiness were purely a matter of how long you'd been awake, that second wind would make no sense. Something else is going on.
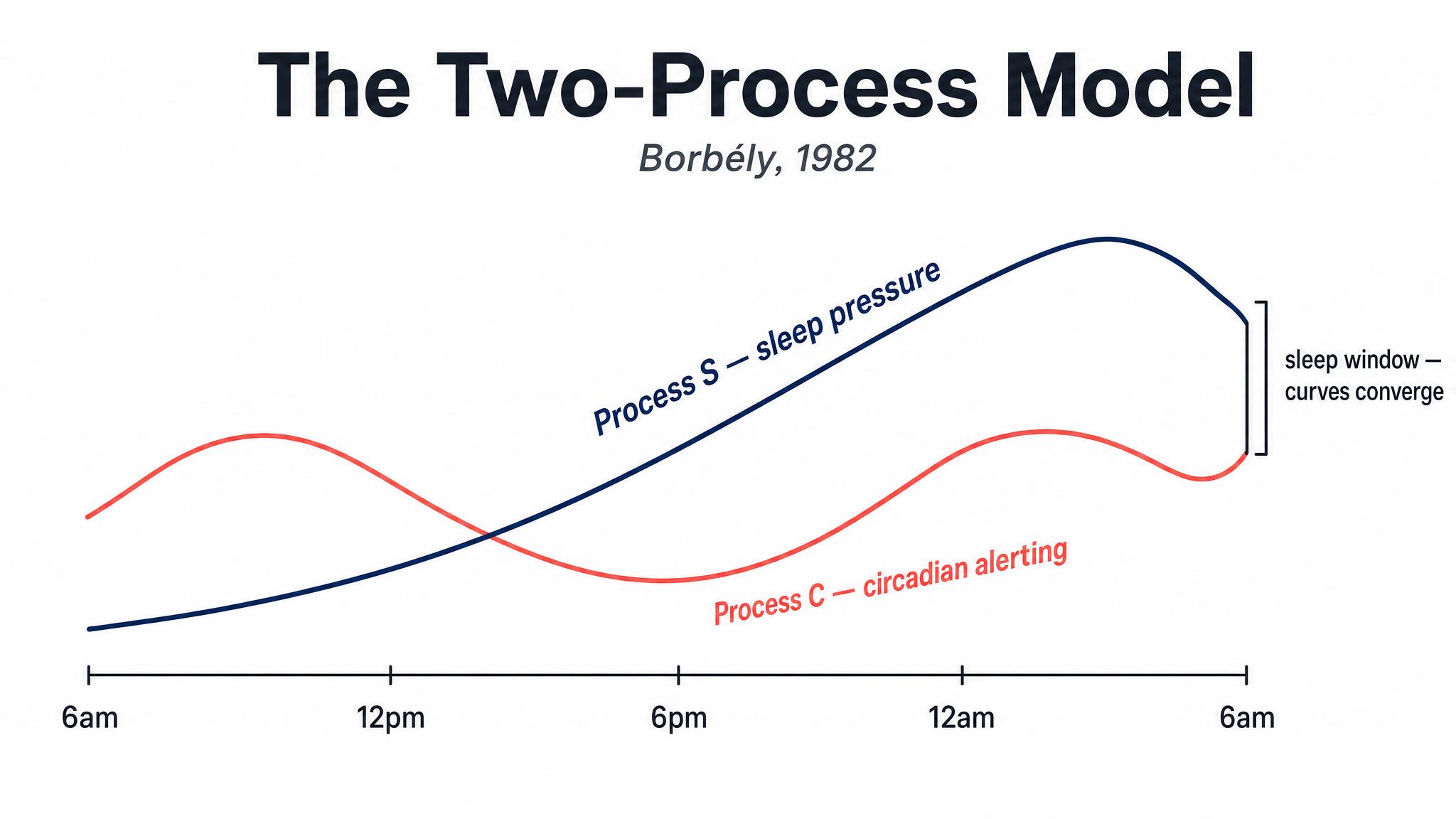
In 1982, the Swiss sleep researcher Alexander Borbély proposed an elegant framework that remains the dominant model of sleep regulation four decades later: the TWO-PROCESS MODEL. It explains sleepiness — and alertness — as the interaction of two independent processes operating simultaneously in your brain.
[short pause]
PROCESS S — S for "sleep-dependent" — is your homeostatic sleep pressure. From the moment you wake up, a chemical called adenosine begins accumulating in your brain as a byproduct of neural metabolism. The longer you stay awake, the more adenosine builds up, and the greater your pressure to sleep becomes. Think of Process S as a slowly rising wave: it starts low when you wake, climbs steadily throughout the day, and reaches its peak after about sixteen hours of continuous wakefulness. When you finally sleep, adenosine is cleared, and Process S drops back down — exponentially at first, then more gradually — resetting the cycle. As Borbély and colleagues described in 2016, if you pull an all-nighter, Process S doesn't plateau — it keeps climbing. That groggy, disoriented, almost painful feeling you experience at four A-M after a night without sleep? That's Process S screaming at you. The pressure has risen far beyond its normal peak, and your brain is increasingly desperate for the restorative clearance that only sleep provides, as Borbély noted in 2022.
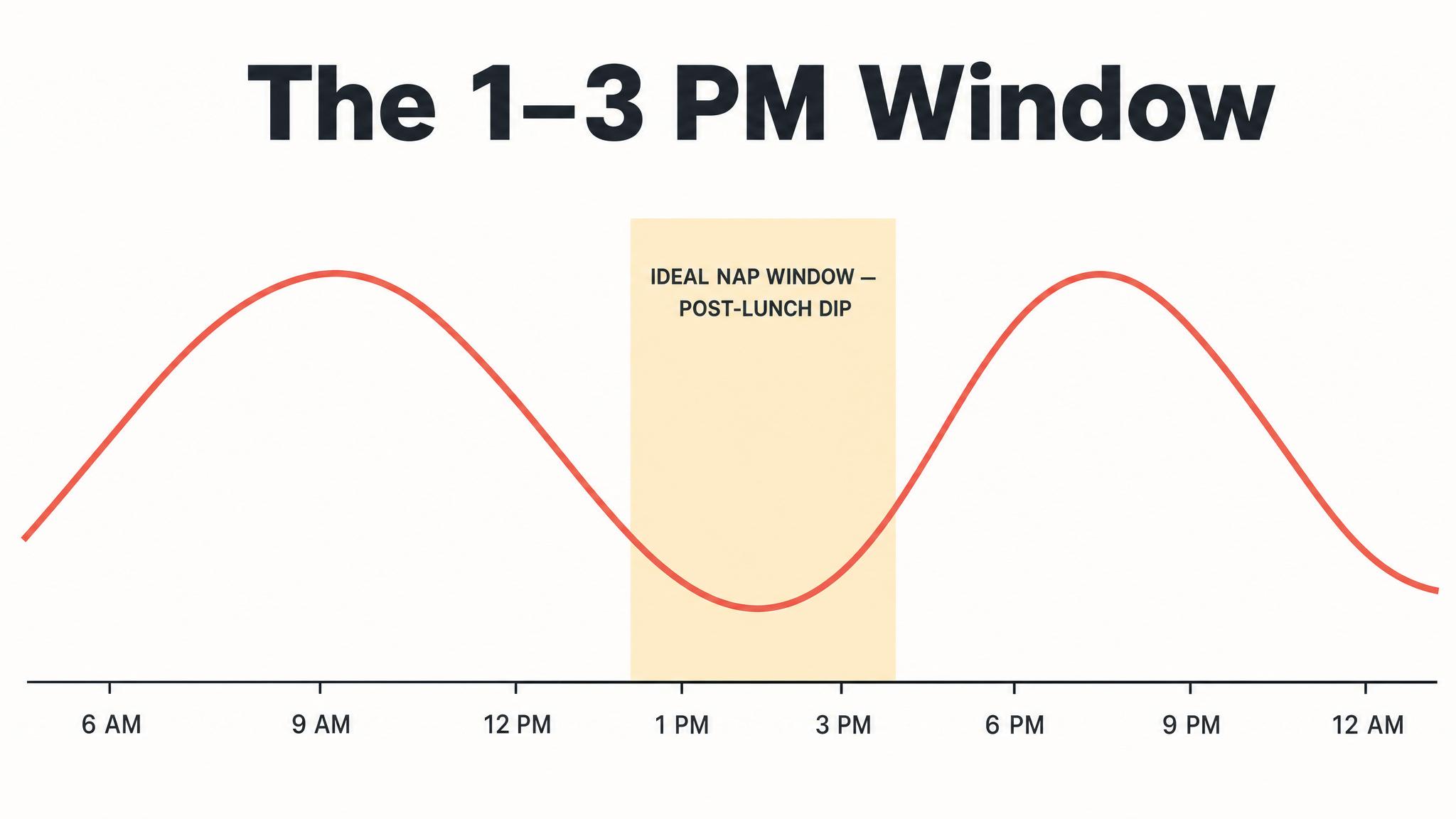
[short pause]
PROCESS C — C for "circadian" — is the oscillating alerting signal generated by your S-C-N. Unlike Process S, which is driven by how long you've been awake, Process C follows its own independent twenty-four-hour rhythm regardless of your behavior. It can be roughly modeled as a sine wave: alerting drive is low in the early morning, rises through the day, dips slightly in the early-to-mid afternoon — that post-lunch sleepiness isn't just about the sandwich — peaks in the early evening, and then drops dramatically as night approaches. As Borbély and colleagues described in 2016, here is the crucial insight: Process S and Process C work in opposition during the day and in concert at night. During your waking hours, the rising sleep pressure of Process S is counteracted by the rising alerting signal of Process C. That's why you don't feel progressively sleepier every single hour — the circadian system is actively propping you up, especially in the late afternoon and early evening. This is also why you can feel a "second wind" around seven or eight P-M; Process C is near its peak alerting force, temporarily masking the high sleep pressure that has accumulated. Then, as evening progresses and Process C plummets, the accumulated Process S pressure is suddenly unopposed. The two curves converge, and you feel powerfully sleepy, as Borbély noted in 2022.
[short pause]
According to the two-process model, what happens if you wake up very early — say, four A-M — but try to stay awake until your normal bedtime? How would Process S and Process C interact differently compared to a normal day? Think through the model: Process S would begin accumulating much earlier in the day, building to extreme levels by evening, while Process C would still follow its normal cycle. The mismatch would create intense sleepiness long before your usual bedtime.
[short pause]
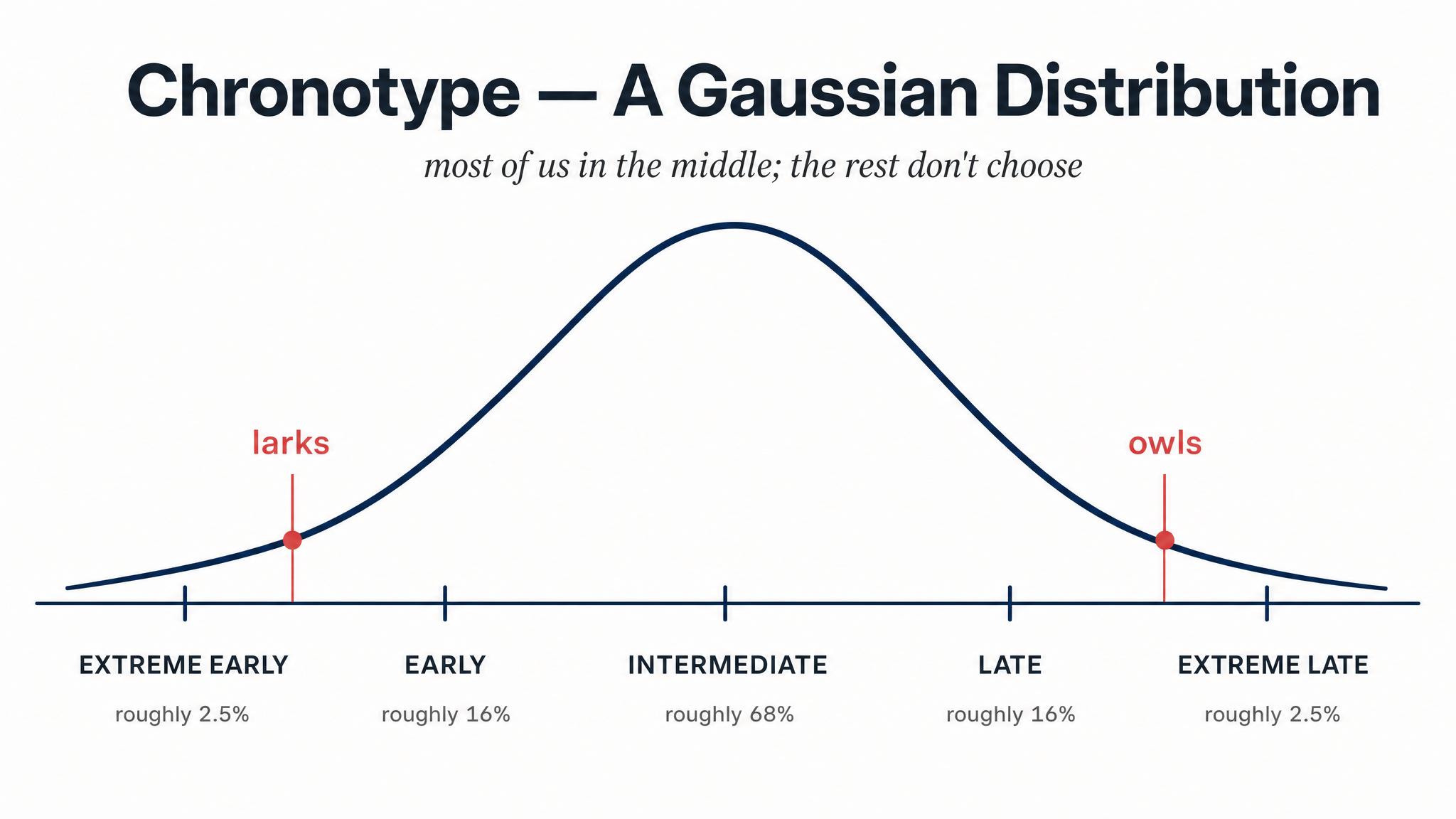
You now know that the circadian clock runs on roughly a twenty-four-hour cycle and that zeitgebers lock it to the external day. But here's a question: is everyone's clock set to the same time? The answer is a definitive no. Just as people vary in height or shoe size, they vary in the timing of their circadian rhythm. This individual variation in preferred sleep-wake timing is called CHRONOTYPE.
Roenneberg, Wirz-Justice, and Merrow in 2003 developed the Munich Chrono-Type Questionnaire, which measures chronotype by asking a deceptively simple question: on free days — when you have no alarm, no obligations, no reason to be anywhere — what time do you naturally fall asleep, and what time do you naturally wake up? The midpoint of that free-day sleep window — say, four A-M for someone sleeping midnight to eight A-M — is your chronotype score. Across tens of thousands of respondents, Roenneberg's team found that chronotype follows a near-Gaussian distribution: most people cluster in the middle, with progressively fewer people at the extreme "lark," meaning early, and "owl," meaning late, ends.
Chronotype is not merely a preference or a lifestyle choice. It has a strong genetic basis, linked to variations in the core clock genes like P-E-R-2, P-E-R-3, and C-R-Y-1. It also shifts dramatically across the lifespan. Children tend to be early types. During adolescence, chronotype delays sharply — teenagers genuinely become more owl-like, not because of poor habits but because of changes in their circadian physiology, including what appears to be a lengthening of the intrinsic circadian period. As Roenneberg and colleagues found in 2003, chronotype reaches its latest point around age twenty, then gradually shifts earlier for the rest of adulthood. This means that when a fifty-five-year-old parent tells their nineteen-year-old child to "just go to bed earlier," they are asking that child to override a biological timing system that is fundamentally different from their own.
[short pause]
Wittmann, Dinich, Merrow, and Roenneberg in 2006 coined the term SOCIAL JETLAG to describe the chronic misalignment between a person's biological clock and the schedule imposed by society. It's calculated simply: the difference between your sleep midpoint on free days and your sleep midpoint on work or school days. If your body naturally sleeps from one A-M to nine A-M — midpoint: five A-M — but your alarm forces you up at six thirty A-M on weekdays — midpoint: roughly three forty-five A-M — you carry about seventy-five minutes of social jetlag. That's the equivalent of flying one time zone west every Friday night and one time zone east every Monday morning, week after week, all semester long.
The consequences are not trivial. Wittmann and colleagues in 2006 found that social jetlag was associated with greater stimulant consumption — particularly caffeine and nicotine — lower psychological well-being, and increased depressive symptoms. The strongest effects appeared in late chronotypes — the owls — because modern school and work schedules overwhelmingly favor early timing. Late types suffer the most misalignment, accumulate the most sleep debt, and show the most pronounced downstream effects.
[short pause]
Calculate your own rough social jetlag. What's the midpoint of your sleep on a typical weeknight versus a typical weekend night? How many minutes separate them? What might this tell you about how well your schedule matches your biology?
[short pause]
If chronotype and social jetlag are real biological phenomena with real consequences, then a prediction follows: changing social schedules to better align with students' biology should produce measurable improvements. In 2018, researchers put this prediction to the test.
Dunster and colleagues in 2018 studied high school students in Seattle before and after the school district delayed start times from seven fifty A-M to eight forty-five A-M. Using wrist-worn actigraphy — objective movement sensors — rather than self-report, they measured students' actual sleep-wake cycles. The results were striking: students gained a median of thirty-four minutes of daily sleep. Crucially, bedtimes did not shift later — students did not simply stay up later to "use" the extra time. Instead, they woke later, sleeping into what their circadian biology was already calling for. The additional sleep was associated with a four point five percent increase in median grades and improved attendance.
This study matters for two reasons. First, it demonstrates that circadian biology has real-world consequences that show up in measurable outcomes like grades and attendance. Second, it illustrates a principle we'll return to throughout this course: sometimes the most powerful intervention is not training people to fight their biology but redesigning schedules to work with it. In the Seattle study, a fifty-five-minute schedule delay translated to thirty-four more minutes of sleep, better grades, and improved attendance — without any change in bedtimes.
[short pause]
As you develop your understanding of sleep science throughout this course, you will encounter bold claims — some well-supported, some overstated. Developing the ability to distinguish between them is a skill this course takes seriously.
Consider a frequently cited claim from Matthew Walker's popular book "Why We Sleep": that sleeping fewer than six hours per night increases your risk of a heart attack by two hundred percent. This statistic has been repeated in media coverage, TED talks, and social media posts millions of times. But what does the underlying evidence actually show? When researchers examine the relevant meta-analyses, the relative risk increase for short sleep and cardiovascular events is real but considerably more modest — typically in the range of twenty to forty-eight percent increased relative risk, depending on the study and the specific outcome measured. And relative risk increases can be misleading without knowing the baseline risk. If your baseline annual risk of a cardiovascular event is one percent, a forty percent relative increase raises it to one point four percent — meaningful at a population level, but quite different from what "two hundred percent increase" implies to a lay reader.
None of this means sleep doesn't matter for cardiovascular health — it clearly does. The point is that scientific literacy requires attending to effect sizes, study designs, and the difference between relative and absolute risk. Throughout this course, when we encounter a claim, we will look at the actual data. This is not skepticism for its own sake — it's the foundation of genuine understanding.
[short pause]
Why might exaggerated health claims about sleep actually backfire? Consider a student who sleeps six hours a night, reads that this "doubles their heart attack risk," and feels overwhelmed and anxious. How might that anxiety itself affect their sleep? We'll explore the relationship between sleep anxiety and insomnia in a later chapter.
[short pause]
Let's review the key insights from this chapter. Your suprachiasmatic nucleus, or S-C-N — a cluster of approximately twenty thousand neurons above the optic chiasm — is the master circadian clock orchestrating twenty-four-hour rhythms in temperature, hormones, alertness, and sleep.
The two-process model explains sleepiness as the interaction of Process S — homeostatic sleep pressure that builds with wakefulness — and Process C — a circadian alerting signal that oscillates independently on a roughly twenty-four-hour cycle.
Zeitgebers, meaning "time givers" — especially light, but also meal timing, exercise, and social cues — entrain your slightly-longer-than-twenty-four-hour internal clock to the external day.
Chronotype is your genetically influenced tendency toward morningness or eveningness. It shifts across the lifespan and follows a normal distribution, with most people intermediate.
Social jetlag — the mismatch between your biological clock and your social schedule — is associated with sleep debt, lower well-being, and increased stimulant use, particularly in late chronotypes.
When school schedules were shifted to align with adolescent biology in the Seattle study, students gained thirty-four minutes of sleep and showed improved grades and attendance.
And finally, good sleep science requires reading claims critically: attend to effect sizes, study designs, and the distinction between relative and absolute risk.
[short pause]
Now that you have the conceptual scaffold — the S-C-N, Process S, Process C, zeitgebers, and chronotype — we're ready to look at what actually happens when you fall asleep. In the next chapter, we'll descend into the architecture of a single night of sleep: the repeating cycles of light sleep, deep slow-wave sleep, and R-E-M sleep, each with distinct neural signatures and distinct biological purposes. You'll learn why the first half of the night looks radically different from the second half, and why this matters for everything from memory to emotional regulation.










The Architecture of a Night — What Happens After You Close Your Eyes
If Chapter 1 was about the 24-hour rhythm that governs when you sleep, Chapter 2 zooms into what happens during the hours you're asleep. Students will discover that sleep is not a uniform state — it is a complex, highly…
If you barely remember your dreams, does that mean you're sleeping badly or sleeping perfectly?
Real talk
Would you rather: Pull an all-nighter before an exam, or sleep 3 hours and wake up feeling like a zombie?
Would you rather
Can you actually 'catch up' on lost sleep over the weekend, or is that just something we tell ourselves?
Debate time
What helps you fall asleep: Total silence, fan/white noise, TV in the background, or music playing?
Quick poll
Forget what doctors say — how many hours of sleep do YOU actually need to feel like a functional human?
Be honest
Hot take: Napping during the day just ruins your sleep at night, so it's never worth it. Agree or disagree?
Unpopular opinion?
Sleep Stage Detective: Live Diagnosis
12-15 minutes1) Instructor calls 3-4 volunteers to front. 2) Each volunteer is secretly given a sleep stage card (N1, N2, N3, or REM) with characteristic behaviors. 3) Volunteers act out their stage simultaneously (N1: slight head nods; N2: stillness with occasional twitches; N3: total stillness, deep breathing; REM: eye movements, facial twitches, body paralysis). 4) Students discuss with immediate neighbors for 2 minutes to identify each stage. 5) Rows call out diagnoses and justify with physiological evidence. 6) Volunteers reveal their stages and class discusses which cues were most diagnostic.
Build Tonight's Sleep: The Hypnogram Challenge
15-20 minutes1) Display blank axes (time vs. sleep depth) on screen. 2) Announce: 'You fall asleep at 11 PM. What happens in the first 90 minutes?' 3) Students confer with neighbors (2 min) to predict the sleep architecture. 4) Call on different sections of theatre: 'Left side—what sleep stage dominates the first cycle? Right side—how long in REM?' 5) Instructor draws the predicted hypnogram based on class input. 6) Advance to next 90-minute cycle. Repeat predictions. 7) After building all 4-5 cycles, overlay the actual typical hypnogram and discuss surprises—why more deep sleep early? Why REM dominates later? 8) Students identify errors in their predictions and explain the physiological reasons.
The Memory Consolidation Tournament
10-12 minutes1) Display two lists of 10 words: List A (emotionally neutral: table, pencil, etc.) and List B (emotionally charged: betrayal, love, fear, etc.). 2) Give students 30 seconds to memorize each list. 3) 2-minute distraction task (solve simple math problems). 4) Students write down all words they remember. 5) Turn to neighbor and compare: Which list had better recall? 6) Take theatre-wide vote: 'More from List A?' vs. 'More from List B?' 7) Discuss results: Why do we remember emotionally charged content better? Connect to REM's role in emotional memory processing and amygdala-hippocampus interaction. 8) Reveal: REM sleep preferentially strengthens emotional memories—they just experienced what happens nightly in REM.
Sleep Cycle Myth Tribunal
15-18 minutes1) Present a controversial sleep claim on screen (e.g., 'You can survive on 4 hours if you only get REM and skip deep sleep' or 'Waking up at exactly 90 minutes is optimal'). 2) Assign theatre sections roles: Front third = Prosecution (argue against), Middle = Defense (argue for), Back = Jury (undecided). 3) Give 3 minutes for sections to discuss with neighbors and gather evidence from lecture concepts. 4) Prosecution presents 2-minute argument using sleep architecture evidence. 5) Defense counters with 2-minute rebuttal. 6) Jury votes using raised hands or phone poll. 7) Instructor debriefs with the science, highlighting which arguments correctly applied concepts about NREM functions, REM necessity, and cycle structure. Repeat with 2nd myth if time allows.
The Slow-Wave Awakening Simulation
8-10 minutes1) Explain: 'We're going to simulate waking someone from different sleep stages.' 2) Show video of someone sleeping. 3) First scenario: Person in N1 (light sleep). Ask class: 'I whisper their name. Do they wake?' Students vote (show of hands). 4) Reveal answer and discuss sleep inertia levels. 5) Second scenario: N3 deep sleep. 'I whisper, then speak loudly, then shake shoulder.' After each escalation, students vote on whether person wakes. 6) Discuss why N3 has high arousal threshold—relate to brain wave activity (delta waves) and physiological restoration functions. 7) Final scenario: REM sleep. 'Despite active brain, would loud noise wake them easily?' Discuss the paradox: active brain but very high awakening threshold. 8) Students turn to neighbor: 'Why does your body essentially paralyze you during REM?' Connect to function and disorders.
Graph Your Night: Sleep Architecture Sketch-Off
12-15 minutes1) Students take out paper/device. 2) Present three scenarios: (a) Normal 8-hour night, (b) Night after 24 hours awake (sleep deprivation), (c) Night after heavy alcohol consumption. 3) For scenario (a), students sketch predicted hypnogram with neighbor (3 min). 4) Random students show sketches on document camera—class votes on most accurate. 5) Reveal correct pattern and discuss. 6) Repeat for scenario (b): 'How would architecture change after sleep deprivation?' Students predict increased N3 early (sleep debt repayment), shortened REM initially. 7) For (c), predict how alcohol suppresses REM and fragments later cycles. 8) Compare all three graphs side-by-side. Debrief: How does the brain prioritize different sleep stages based on physiological need?
Transcript
Imagine two students, Maya and Jake, both getting exactly six hours of sleep before an exam. Maya goes to bed at midnight and sets her alarm for six A-M. Jake stays up until three A-M and sleeps until nine A-M. They sleep for the same duration, but Maya wakes up feeling foggy and emotionally raw, while Jake feels physically sluggish but mentally sharp. Same total hours, radically different outcomes. Why?
The answer lies in a fact that surprises most people: sleep is not one thing. It is a structured sequence of biologically distinct stages, each performing different work, and the balance between those stages shifts dramatically across the night. Cut from one end, and you lose different capacities than if you cut from the other. Understanding this architecture, this hidden blueprint of a night, changes how you think about every sleep decision you make.
[short pause]
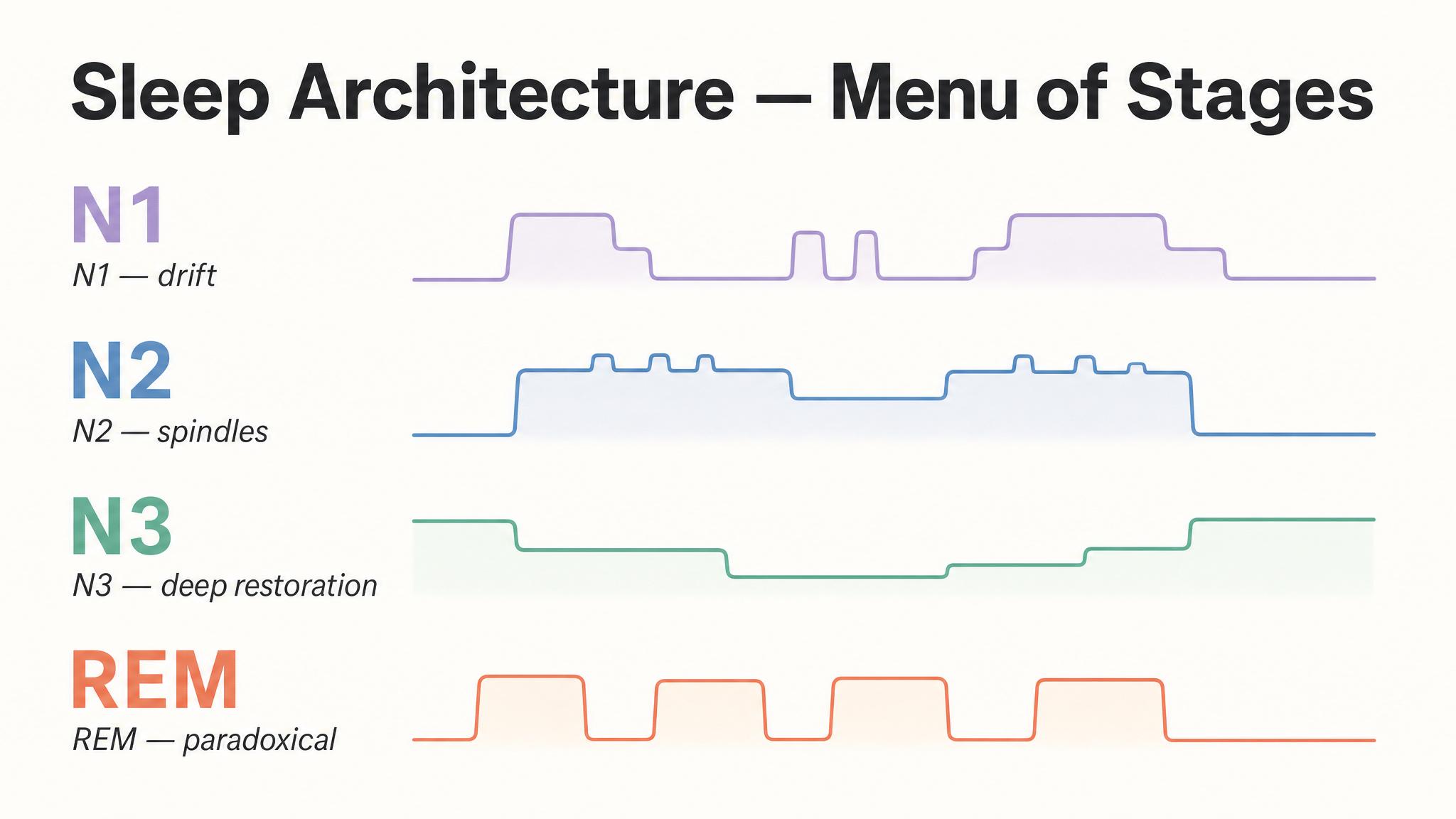
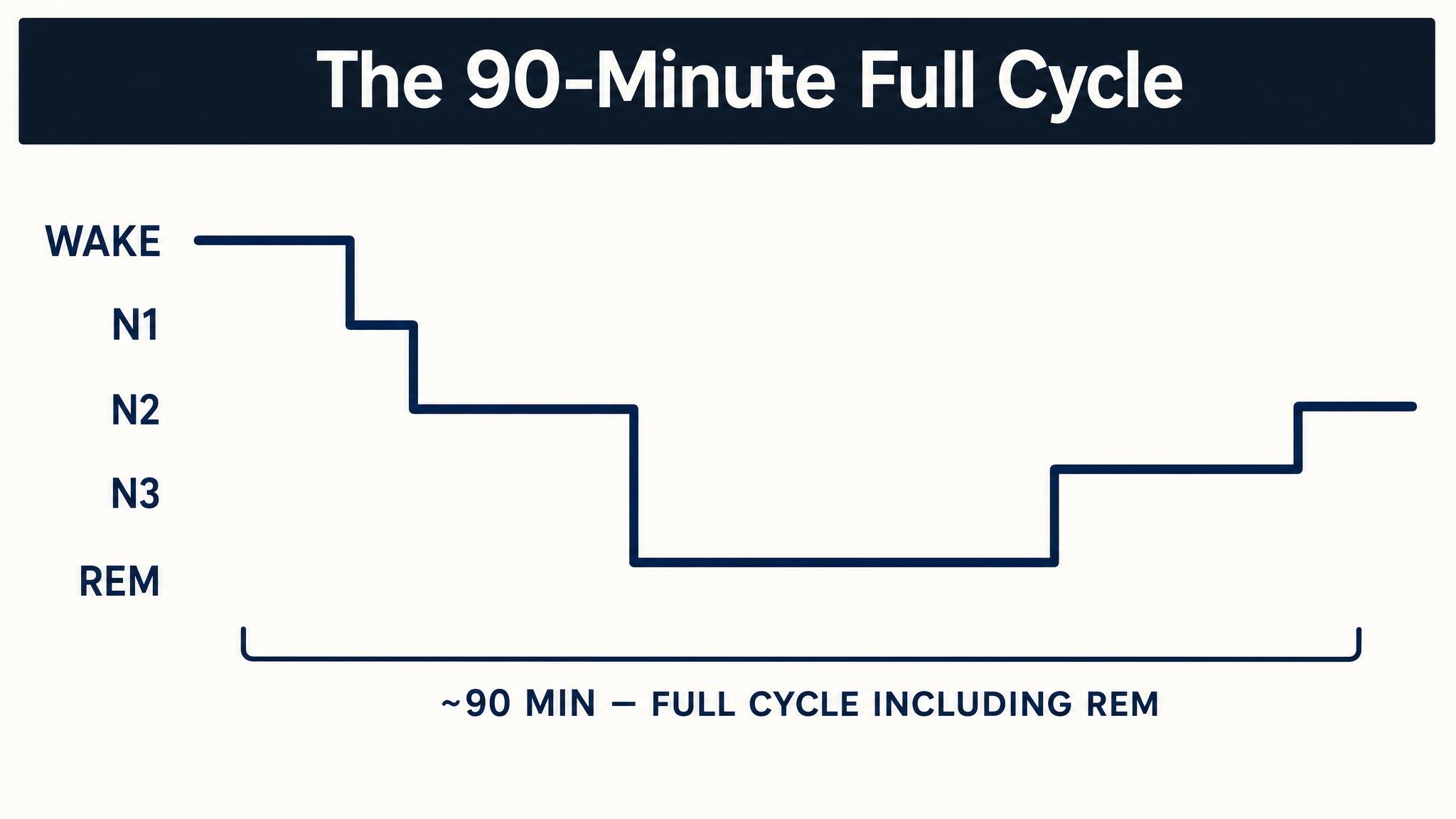
When you close your eyes and drift off, your brain does not simply power down like a laptop entering standby mode. Instead, it begins executing a complex, highly organized sequence of neural states that cycle roughly every ninety minutes throughout the night. This recurring pattern is called the ULTRADIAN RHYTHM, a within-sleep rhythm nested inside the twenty-four-hour circadian rhythm you studied in the previous lesson.
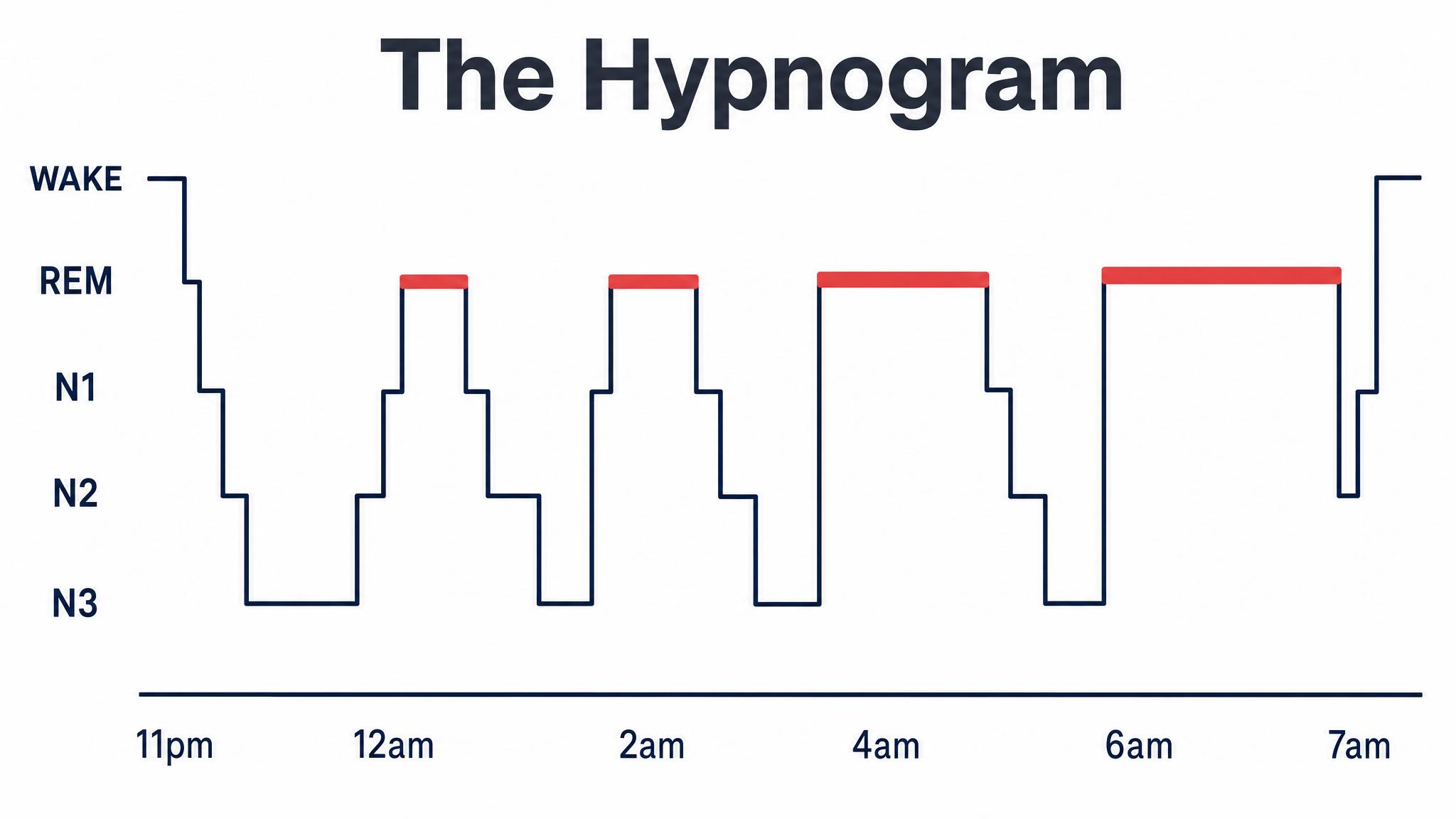
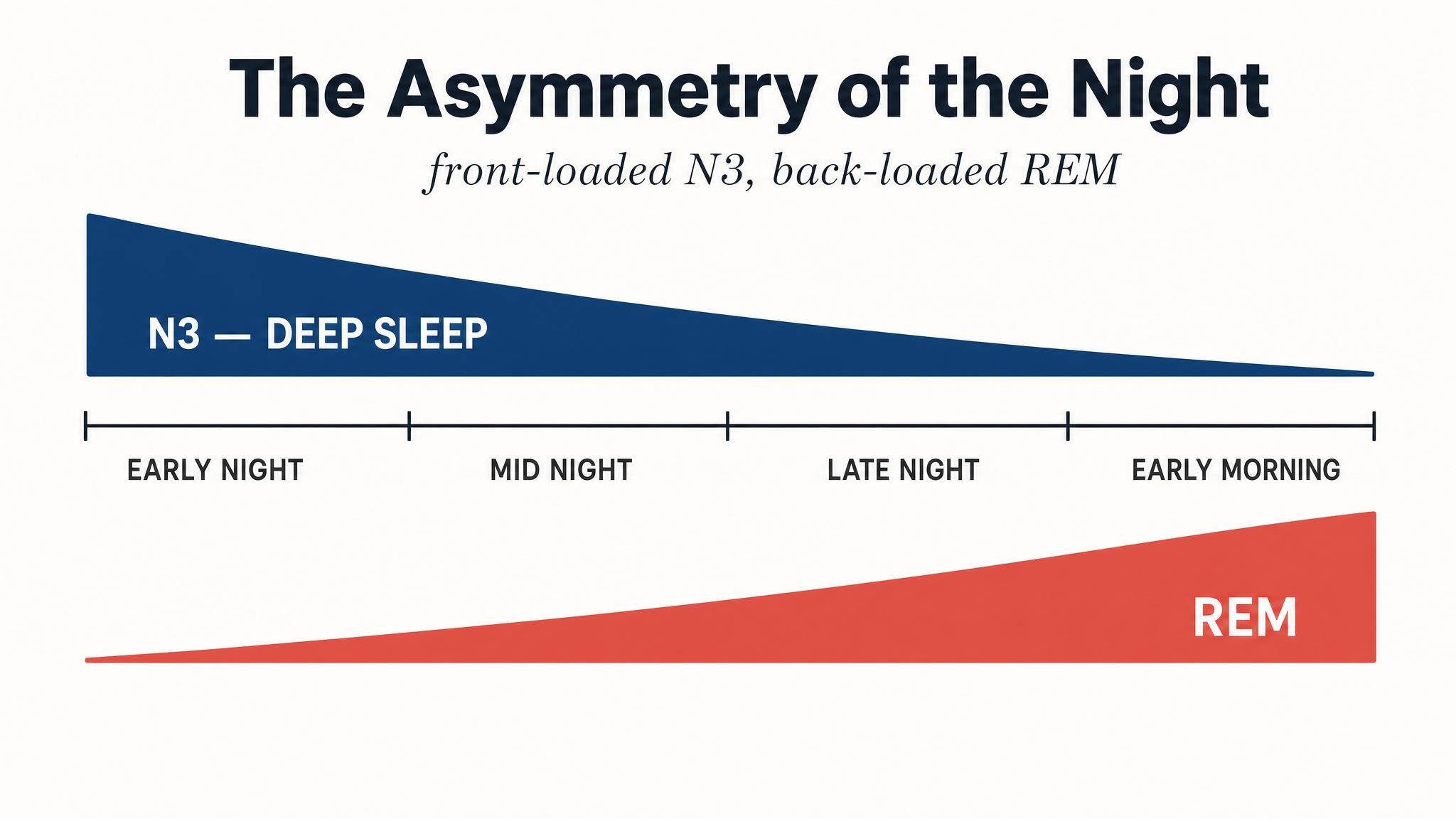
Sleep scientists divide this sequence into two broad categories: non-rapid eye movement sleep, which itself has three stages called N-one, N-two, and N-three, and rapid eye movement sleep, the stage most associated with vivid dreaming. A single pass through all stages constitutes one sleep cycle, and a typical eight-hour night contains four to six of these cycles, as Patel and colleagues reported in twenty twenty-four. But here is the critical insight that will frame this entire chapter: the cycles are not identical copies. Early cycles are dominated by deep non-rapid eye movement sleep; later cycles are dominated by rapid eye movement sleep. This asymmetry has profound consequences.
[short pause]
The lightest stage of sleep, N-one, is the brief transition between wakefulness and sleep, typically lasting only one to five minutes. Your brain shifts from the alpha waves of relaxed wakefulness to the slower theta waves of early sleep. Muscles begin to relax, and you may experience hypnagogic phenomena: fleeting sensory hallucinations, the sensation of falling, or sudden whole-body muscle jerks called hypnic myoclonia, as Patel and colleagues described.
N-one is so light that most people, if awakened from it, will deny having been asleep at all. It constitutes only about five percent of total sleep time and is not considered restorative in itself. Think of it as a doorway, not a room you spend much time in, but one you must pass through repeatedly as you transition between deeper stages across the night.
[short pause]
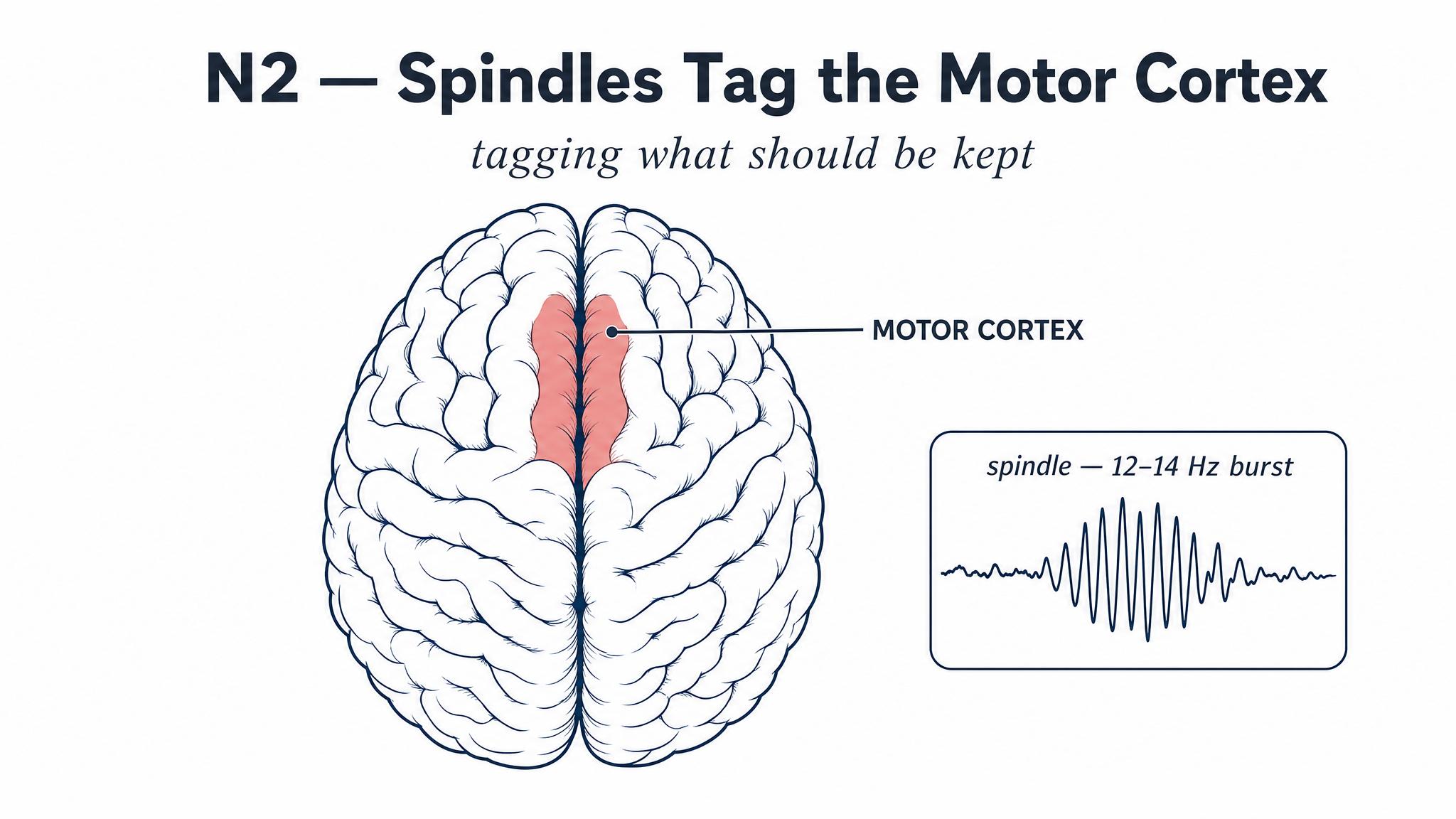
You spend more time in N-two than any other stage, roughly forty-five to fifty-five percent of total sleep time. And while it has historically been treated as a mere waypoint between light and deep sleep, recent research has revealed N-two to be a remarkably active processing stage in its own right, thanks to two distinctive electrical signatures: SLEEP SPINDLES and K-COMPLEXES.
Sleep spindles are brief bursts of rhythmic neural activity, eleven to sixteen hertz, generated by the thalamus. They appear as tight, waxing-and-waning oscillations on an E-E-G readout, and they are far more than electrical noise. In a landmark study, Nishida and Walker demonstrated in two thousand and seven that improvements in motor skill memory, the kind of memory involved in learning a piano sequence or a tennis serve, correlated specifically with the density of sleep spindles during N-two sleep. More strikingly, these spindles were regionally specific: if a participant learned a right-hand motor task, spindle density increased over the left motor cortex, the exact hemisphere controlling that hand. The brain was not just passively resting; it was rehearsing the day's motor learning at an anatomically precise location.
K-complexes, the other hallmark of N-two, are large, sharp waveforms that appear to serve a gating function. When an external sound occurs during N-two sleep, a door closing, a car horn, a K-complex fires in response, essentially evaluating whether the stimulus warrants waking up. If the sound is deemed non-threatening, sleep continues. K-complexes thus function as a night-watch system, filtering irrelevant noise while protecting the continuity of sleep, according to Patel and colleagues.
[short pause]
Consider this: if N-two sleep spindles are critical for motor memory consolidation, what would you predict happens to a musician's performance after a night of fragmented sleep where N-two is repeatedly interrupted, even if total sleep duration is preserved?
[short pause]
N-three, also called slow-wave sleep, is the deepest stage of non-rapid eye movement sleep, and, in many ways, the most physiologically powerful. It is characterized by high-amplitude, low-frequency DELTA WAVES, zero-point-five to two hertz, that sweep across the cortex in slow, synchronized oscillations. This is the stage from which you are hardest to wake, and the stage in which your brain and body perform some of their most critical maintenance work.
Physiologically, N-three is when the body releases the majority of its daily growth hormone, driving tissue repair, muscle recovery, and immune system strengthening, as Patel and colleagues noted. This is why athletes who lose deep sleep see measurable declines in physical recovery, a connection we will explore further in a later chapter.
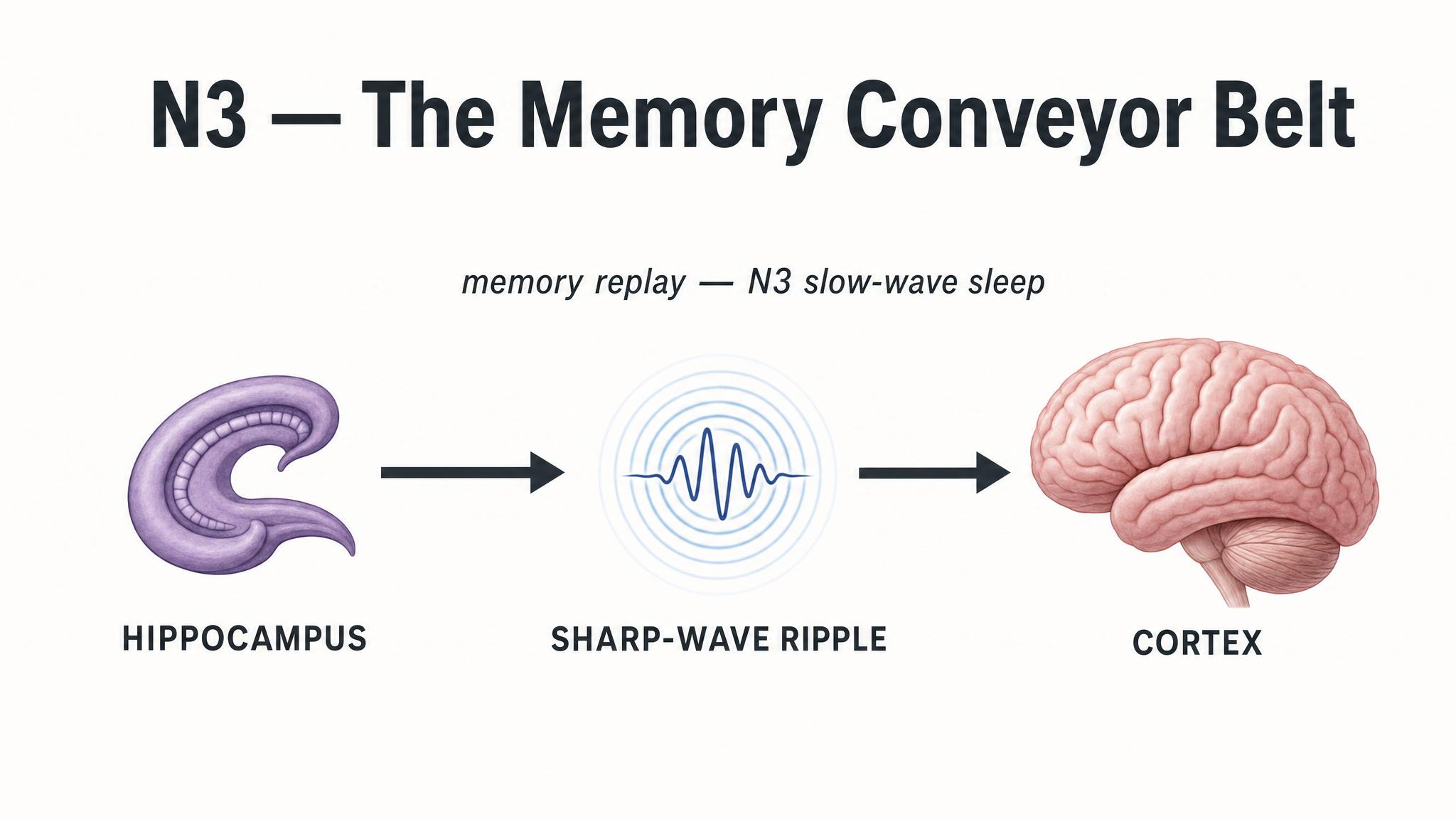
But N-three's contributions extend far beyond the body. The ACTIVE SYSTEM CONSOLIDATION model, proposed by Diekelmann and Born in twenty ten, describes a remarkable neural process that unfolds during slow-wave sleep. Here is how it works: memories that were initially encoded in the hippocampus during the day are spontaneously reactivated during slow-wave sleep. These reactivations occur during sharp wave-ripples in the hippocampus, which are temporally coordinated with thalamic sleep spindles and the slow cortical oscillations that define N-three. This triple-nested coordination, ripple within spindle within slow oscillation, creates a neural conveyor belt that gradually transfers memory representations from the hippocampus to more stable, long-term storage sites in the neocortex, as Rasch and Born described in twenty thirteen.
This process particularly benefits DECLARATIVE MEMORIES, memories for facts, events, and explicit knowledge. In experimental paradigms, participants who studied word pairs or spatial layouts before sleep showed significantly better recall after sleep rich in slow-wave activity, compared to equivalent periods of wakefulness, according to findings by Diekelmann and colleagues in two thousand and nine. The effect was especially strong for weakly encoded items, precisely the kind of fragile new knowledge that needs consolidation most.
[short pause]
As Diekelmann and Born noted in twenty ten, quote, "The sleeping brain is not simply maintaining memories, it is actively reorganizing them, extracting their gist, and integrating them into existing knowledge networks."
[short pause]
More recently, researchers have linked N-three to the GLYMPHATIC SYSTEM, a waste-clearance mechanism in which cerebrospinal fluid flushes through brain tissue, clearing metabolic byproducts including amyloid-beta, a protein implicated in Alzheimer's disease. While this research is still emerging and the precise stage-specificity is under active investigation, the current evidence suggests that deep non-rapid eye movement sleep provides the optimal conditions for this neural housekeeping.
[short pause]
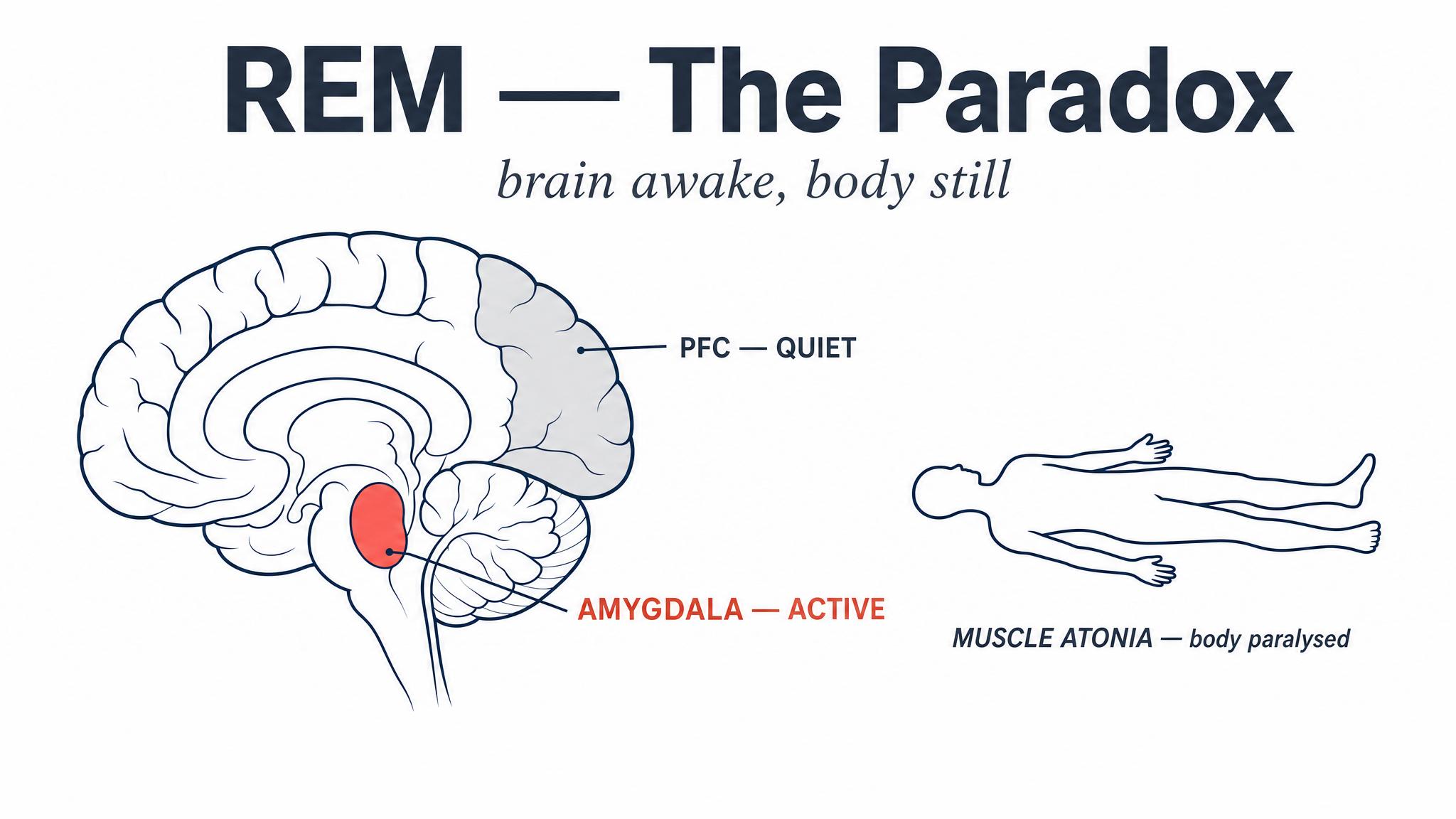
After roughly seventy to ninety minutes of progressively deepening non-rapid eye movement sleep, something extraordinary happens. The brain reverses course and enters rapid eye movement sleep, a stage so physiologically distinct from everything that came before that its discoverers initially called it "paradoxical sleep." The paradox: your brain becomes almost as electrically active as when you are awake, yet your body becomes almost completely paralyzed.
During rapid eye movement sleep, the brainstem sends inhibitory signals to motor neurons throughout the body, producing a state called muscle atonia. Your voluntary muscles are effectively disconnected. This paralysis appears to be a protective mechanism, preventing you from physically acting out the vivid, story-like dreams that characterize this stage. When the atonia mechanism fails, the result is rapid eye movement sleep behavior disorder, in which people kick, punch, or leap from bed during dreams.
Meanwhile, the brain is extraordinarily busy. The emotional centers, particularly the amygdala and the anterior cingulate cortex, become highly active, while the prefrontal cortex, responsible for logical oversight and impulse control, becomes relatively quiet. This unique neurochemical environment, high in acetylcholine, low in norepinephrine and serotonin, appears to create ideal conditions for emotional memory processing.
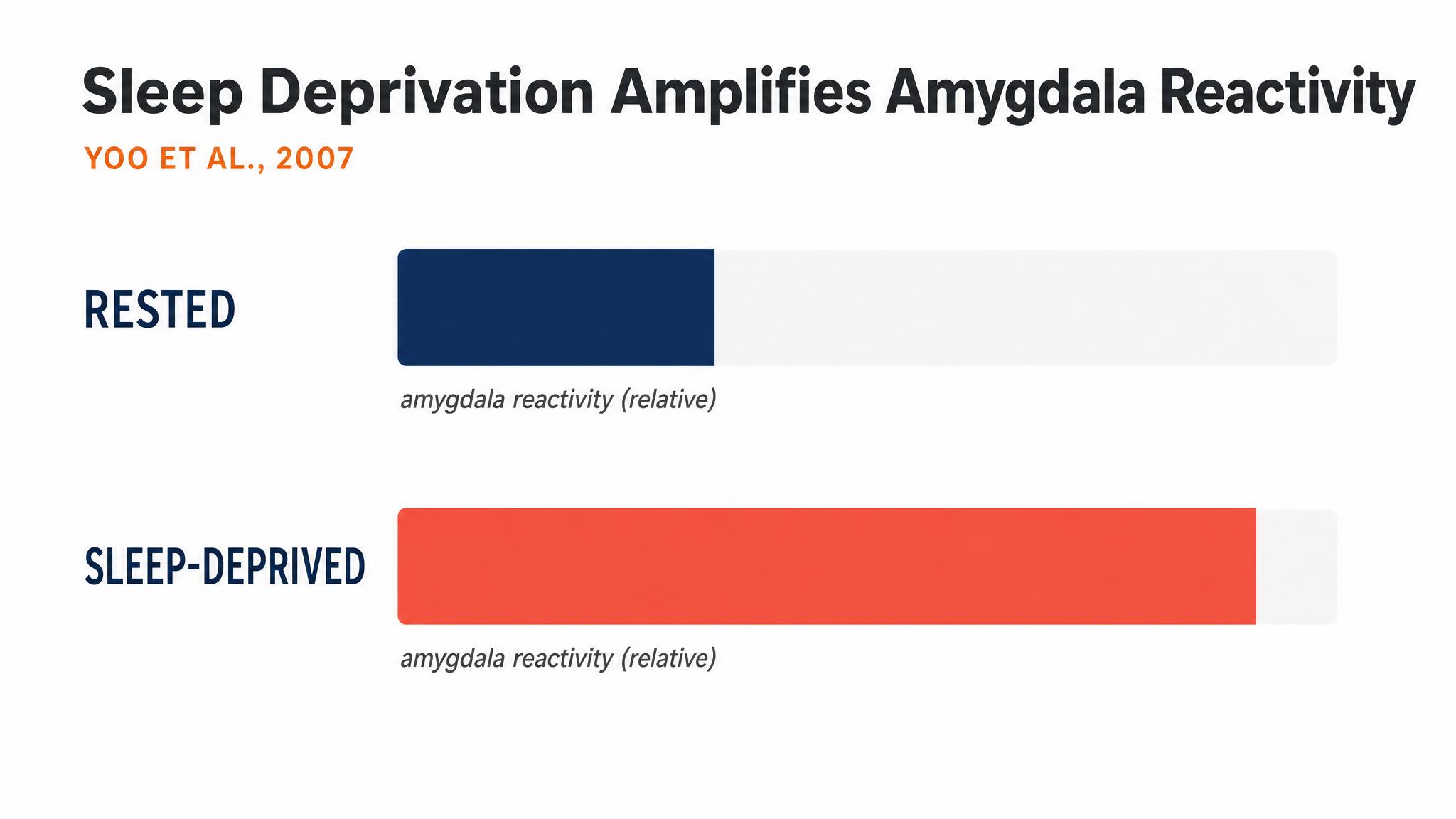
Goldstein and Walker demonstrated in twenty fourteen that sleep deprivation amplified amygdala reactivity to negative emotional stimuli by roughly sixty percent, accompanied by reduced connectivity between the amygdala and the medial prefrontal cortex, the neural circuit that normally keeps emotional responses in check. Their findings suggest that sleep, and rapid eye movement sleep in particular, plays a role in recalibrating emotional responses overnight.
[short pause]
It is tempting to assign rapid eye movement sleep sole responsibility for emotional regulation, and some popular accounts do exactly that, claiming that without sufficient rapid eye movement sleep, you become essentially emotionally irrational. The evidence, however, is more nuanced than this narrative suggests.
Tempesta and colleagues conducted a comprehensive review in twenty eighteen of sleep and emotional processing, concluding that while evidence converges on rapid eye movement sleep benefiting emotional processing, quote, "contradictory findings exist and crucial aspects remain unclear." Total sleep deprivation studies consistently show emotional dysregulation, but isolating the specific contribution of rapid eye movement sleep versus non-rapid eye movement sleep remains methodologically challenging. Some emotional memory benefits appear even after short naps dominated by non-rapid eye movement sleep, according to Diekelmann and colleagues.
Schönauer and Rasch proposed in twenty fifteen a more integrated model: non-rapid eye movement slow-wave sleep initially consolidates the memory content of emotional experiences, while rapid eye movement sleep subsequently integrates those consolidated traces into broader neocortical networks, gradually decontextualizing them, separating the emotional charge from the factual content. This two-stage model is elegant, but it also makes clear that no single stage works alone. The lesson for students of sleep science is important: be cautious of any claim that assigns a complex psychological function to a single sleep stage. Sleep stages work as a coordinated system.
[short pause]
Consider this distinction: "rapid eye movement sleep is associated with emotional memory processing" versus "rapid eye movement sleep is solely responsible for emotional regulation." What kind of evidence would you need to move from the first claim to the second? Why is that evidence so difficult to obtain?
[short pause]
Now that you understand what each stage does individually, we can examine how they combine across a full night, and why the sequence matters as much as the stages themselves.
A typical night of eight hours contains four to six complete sleep cycles, each lasting approximately ninety minutes. But these cycles are not interchangeable. The first two cycles, roughly the first three hours of sleep, are dominated by deep N-three slow-wave sleep, with only brief periods of rapid eye movement sleep. As the night progresses, N-three diminishes and rapid eye movement periods grow progressively longer. By the final cycle, typically between six and eight A-M for someone who fell asleep around eleven P-M, rapid eye movement sleep may last forty to sixty minutes, while N-three has nearly vanished, as Patel and colleagues described.
This asymmetry is not random. It is driven by the two regulatory processes you learned about in the previous lesson. Process S, homeostatic sleep pressure, primarily adenosine accumulation, is at its peak when you first fall asleep, and it is this intense pressure that drives the brain down into deep N-three sleep early in the night. As N-three sleep dissipates adenosine, Process S weakens. Meanwhile, Process C, the circadian signal, increasingly favors rapid eye movement sleep as the night progresses, with rapid eye movement propensity peaking in the early morning hours. The two processes together sculpt the shifting landscape of your sleep cycles.
This brings us back to Maya and Jake from our opening scenario. Maya, sleeping from midnight to six A-M, captured most of her deep N-three sleep, front-loaded in the first three hours, but lost her final, rapid eye movement-rich cycles. Jake, sleeping from three A-M to nine A-M, slept through the circadian rapid eye movement window but missed the initial deep sleep surge, though his high sleep pressure would have partially compensated by driving rapid descent into N-three. The result: Maya preserved physical restoration but sacrificed emotional processing; Jake got more rapid eye movement sleep but less total deep sleep. Neither got a complete night's architecture.
[short pause]
Based on the front-loading of N-three and back-loading of rapid eye movement sleep, predict which functions would be most impaired by consistently waking up two hours earlier than your body wants. Now consider: which functions would be most impaired by consistently going to bed two hours later?
[short pause]
Understanding sleep architecture transforms abstract neuroscience into practical decision-making. Here are three implications that will recur throughout this course.
First, caffeine timing reshapes your stages. Caffeine blocks adenosine receptors, the very mechanism that drives Process S and the descent into deep N-three sleep. Consuming caffeine in the afternoon or evening does not merely delay sleep onset; it selectively reduces deep slow-wave sleep even when total sleep duration is preserved. You will explore this mechanism in detail in a later chapter, but the architectural logic should already be clear: anything that blunts adenosine's signal will disproportionately erode the N-three-rich early cycles.
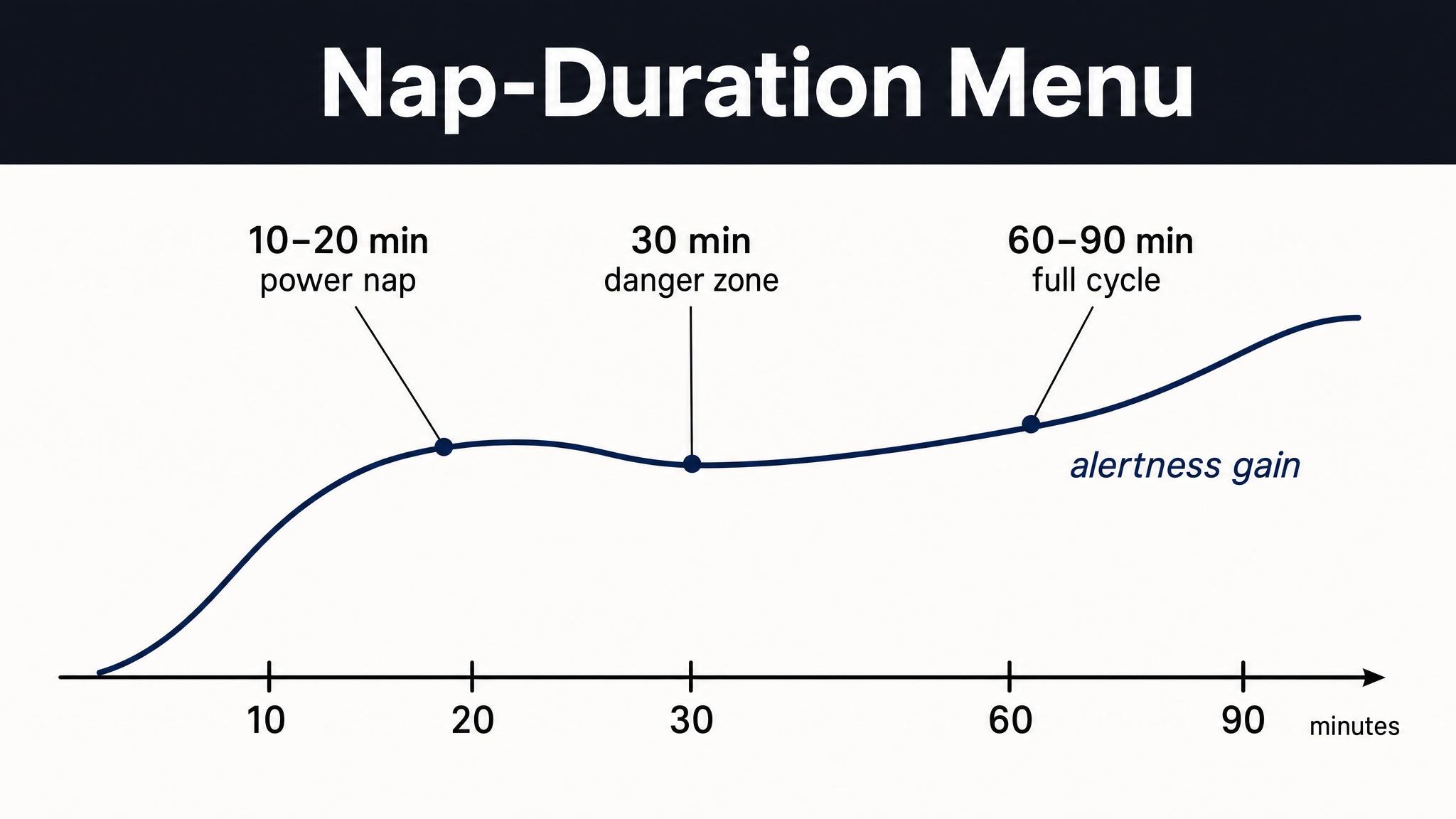
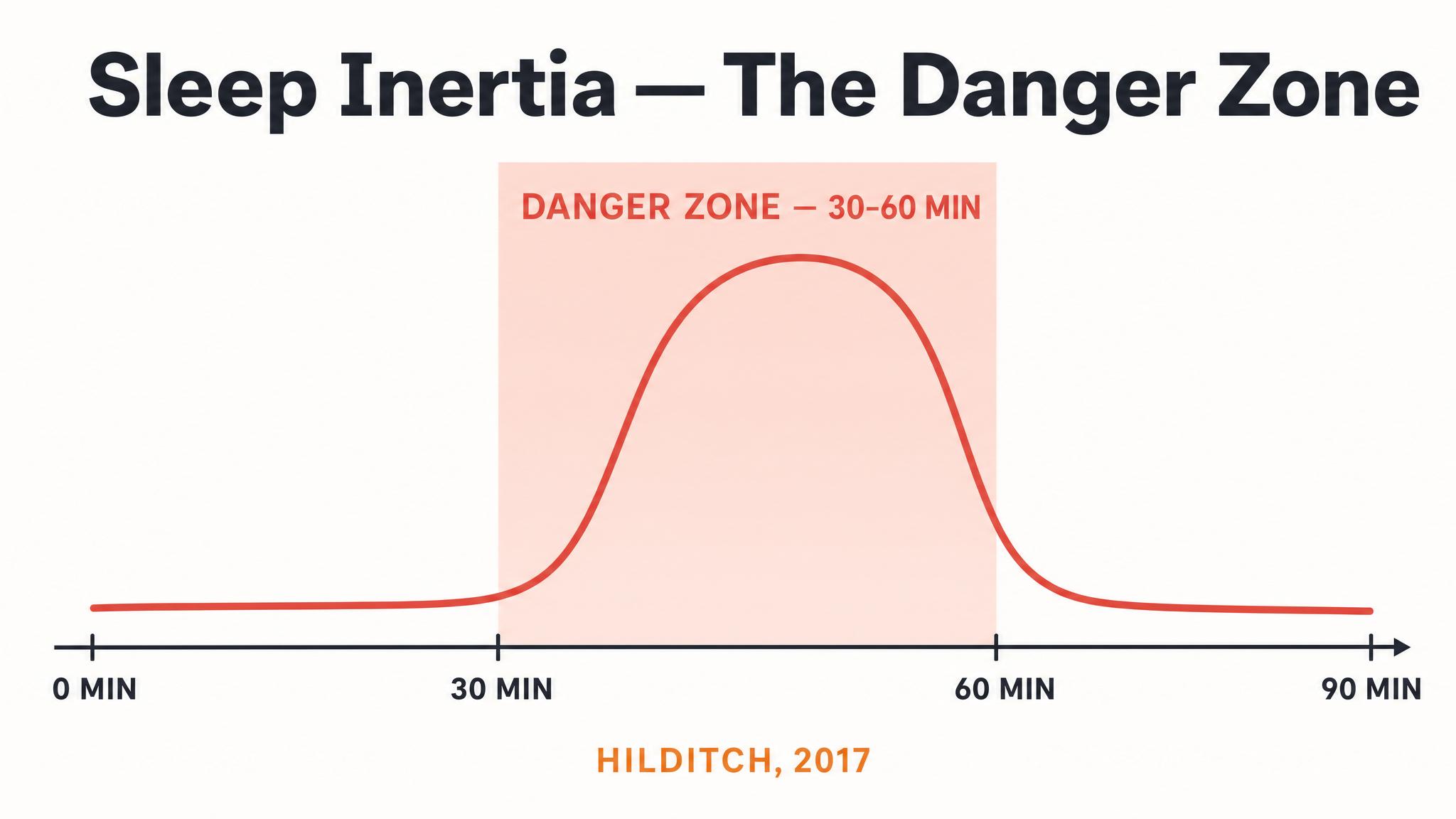
Second, nap design depends on which stage you need. A twenty-minute nap captures mostly N-two, gaining you spindle-mediated motor consolidation and mild restoration. A ninety-minute nap allows a complete cycle including N-three and possibly rapid eye movement sleep, offering deeper restoration and broader memory benefits, but also risks sleep inertia upon waking. When you study nap architecture in a later chapter, you will design naps based on which stage's functions you most need at that moment.
Third, exercise timing interacts with deep sleep. Vigorous exercise increases N-three sleep, but only when performed with appropriate timing relative to circadian phase. Evening exercise too close to bedtime can elevate core body temperature and delay sleep onset, paradoxically reducing the N-three it would otherwise enhance. A later chapter will explore the optimal timing window.
[short pause]
In the previous lesson, you learned that Process S, homeostatic sleep pressure, and Process C, circadian alerting signal, jointly determine when you sleep. Now you can see that they also determine how you sleep. Process S, driven by adenosine accumulation, powers the deep slow-wave sleep that dominates early cycles. As N-three discharges that pressure, Process C's circadian gate swings open for rapid eye movement sleep. The two processes are not just scheduling sleep; they are programming its internal structure.
This interconnection means that anything disrupting Process S or Process C does not just change sleep timing; it changes sleep composition. Shift workers, jet-lagged travelers, and students pulling all-nighters do not merely sleep fewer hours, they sleep differently configured hours, with potentially altered ratios of N-three to rapid eye movement sleep. The architectural lens makes visible what a simple "hours slept" metric cannot.
[short pause]
Let's review the key takeaways. Sleep is not a uniform state but a structured sequence of distinct stages, N-one, N-two, N-three, and rapid eye movement sleep, cycling roughly every ninety minutes in an ultradian rhythm.
N-two sleep spindles consolidate motor memories at regionally specific brain locations, while K-complexes filter external stimuli to protect sleep continuity.
N-three, slow-wave sleep, is critical for growth hormone release, immune function, glymphatic waste clearance, and the active consolidation of declarative memories through coordinated hippocampal replay.
Rapid eye movement sleep is associated with emotional memory processing and affective recalibration, but its specific contribution is more nuanced than popular claims suggest. Non-rapid eye movement and rapid eye movement stages work as a coordinated system.
Sleep architecture shifts across the night: N-three dominates the first half, rapid eye movement sleep dominates the second. This means truncating sleep from different ends disproportionately costs different functions.
Process S drives early deep sleep; Process C gates later rapid eye movement sleep. The two-process model from the previous lesson directly programs the internal structure of each night.
And finally, practical decisions about caffeine, nap timing, exercise, and sleep schedules all reshape sleep architecture by selectively affecting specific stages.
[short pause]
Now that you understand the architecture of a single night, the next chapter will ask: what happens when this architecture breaks down? We will examine the neuroscience and health consequences of sleep deprivation, both total and partial, and explore why even modest sleep restriction, accumulated over consecutive nights, produces cognitive and physiological deficits that most people fail to notice in themselves. You will learn about sleep debt, the myth of "catching up" on weekends, and the dose-response relationship between lost sleep and impaired performance.








Light as a Drug — The Most Powerful Button on Your Clock
Having established what the circadian clock is (Chapter 1) and what it orchestrates during sleep (Chapter 2), this chapter reveals the single most powerful tool for controlling that clock: light. Students will learn…
Do you think 'I'm just naturally a night owl' is a biological fact or an excuse people make? Turn to someone and argue about it.
Debate time
Would you rather: give up ALL screens after 8pm for a month OR wake up every morning to a slowly brightening alarm light (no snooze button)?
Would you rather
When you're scrolling your phone in bed in the dark, do you actually believe it's messing with your sleep or do you think that's overblown?
Honest question
If fixing your sleep meant giving up ONE thing for a week — coffee, late-night screens, or sleeping in — which would you sacrifice?
Real talk
Someone turns on full overhead lights at a 2am house party. Are they a villain or doing everyone a favor?
Hot take
Your roommate leaves a bright desk lamp on all night while they study. Are you allowed to be mad about it?
Roommate drama
Your Light Exposure Autopsy
12-15 minutesStudents create a 24-hour timeline of their light exposure yesterday on paper, marking approximate times and types of light (sunlight, screens, indoor lights). They then turn to 1-2 neighbors and compare timelines, identifying: (1) Who got morning light earliest? (2) Who had the most evening blue light? (3) Whose circadian system got the clearest 'day/night' signal? Groups call out their findings to the class, and instructor maps patterns on board showing common circadian sabotage patterns (late morning light, excessive evening exposure). Concludes with students circling their #1 modifiable behavior.
Phase-Response Curve Live Simulation
10-12 minutesInstructor presents a scenario: 'You're jet-lagged, trying to shift your clock 3 hours earlier.' Projects a phase-response curve on screen. Students vote (by raising hands or using poll) on whether light at different times (6am, 10am, 2pm, 8pm, 11pm) will advance, delay, or have no effect on their clock. After each vote, instructor reveals answer using the curve and explains. Students track points for correct answers. Final round: instructor gives complex scenario (shift worker, international traveler) and students must recommend light timing. Winning student/row gets recognition.
Blue Light Courtroom Debate
15-18 minutesDivide room into three sections: 'Prosecution' (left side), 'Defense' (right side), 'Jury' (middle). Present case: 'Blue-blocking glasses and night mode are sufficient for healthy evening screen use.' Prosecution argues why evening blue light is harmful despite these tools (melanopsin sensitivity, intensity still matters, behavioral cues). Defense argues tools are adequate (spectral filtering, practicality, dose-response). Each side has 3 minutes to consult with neighbors and build argument. Selected students from each side present (2 min each). Jury votes. Instructor debriefs with evidence on lux thresholds, ipRGC spectral sensitivity, and practical recommendations.
Lux Detective Challenge
8-10 minutesInstructor shows photos of 6 different environments (bedroom at night with phone, outdoor shade, cloudy day, office with windows, direct sunlight, dim restaurant). Students write down estimated lux levels for each (working with neighbor for 4 minutes). Instructor then reveals actual values and explains: outdoor shade (~1,000 lux) vs. bright indoor (~500 lux) vs. sunset (~10-50 lux) vs. screens (~50-100 lux). Students calculate how close they were. Discussion focuses on why we wildly underestimate how bright 'outdoors' is versus 'indoors' and why '10 minutes outside' beats '2 hours by office window' for circadian entrainment.
Morning Light Protocol Design Sprint
12-15 minutesPresent 4 different personas on screen (shift worker, college student with 8am class, new parent, remote worker). Students pair with neighbor, select one persona, and design a realistic morning light protocol: What time? What duration? What intensity? Indoor/outdoor? Backup plan? (6 minutes). Then use 'gallery walk alternative': each pair shares their protocol with the pair in front/behind them (3 minutes). Instructor cold-calls 4-5 pairs to share their best protocol. Class votes on most practical. Instructor provides expert feedback on each, highlighting melanopsin sensitivity, minimum effective dose (~20-30 min at 1000+ lux), and timing relative to wake.
Circadian Sabotage Autopsy
10-12 minutesInstructor presents 3 real case studies of people with sleep/energy problems (anonymized or fictional): descriptions include full light exposure patterns. Students work with neighbors to diagnose which aspect of light exposure is most problematic for each case using melanopsin/ipRGC concepts (e.g., late morning light → delayed phase; bright evening light → can't fall asleep; insufficient daytime light → weak circadian amplitude). Each group writes their diagnosis and #1 recommended intervention. Instructor randomly calls on students to share diagnoses. Class discovers multiple people identified different issues—instructor uses this to show how light affects circadian system through multiple mechanisms (phase shifting, amplitude, direct alertness via ipRGCs).
Transcript
In 2013, Kenneth Wright took eight volunteers camping in the Colorado Rockies for a week. No flashlights. No phones. No electric light of any kind. After just seven days under natural skies, something remarkable happened: every participant's internal clock shifted roughly two hours earlier, their melatonin began rising right at sunset, and perhaps most striking, the large differences in sleep timing that had separated night owls from morning larks shrank dramatically. This finding, reported by Wright and colleagues in 2013, came from a simple intervention. The campers hadn't taken a sleeping pill, adopted a mindfulness practice, or forced themselves into bed earlier. They had simply changed their light environment.
[short pause]
That experiment crystallizes the central lesson of this chapter. If Chapter One introduced your circadian clock and Chapter Two showed you what that clock orchestrates during sleep, this chapter reveals the single most powerful input to that clock: LIGHT. Not light as you experience it visually — not color, not images, not beauty — but light as a raw signal of time, measured by cells in your eye that you never knew you had.
[short pause]
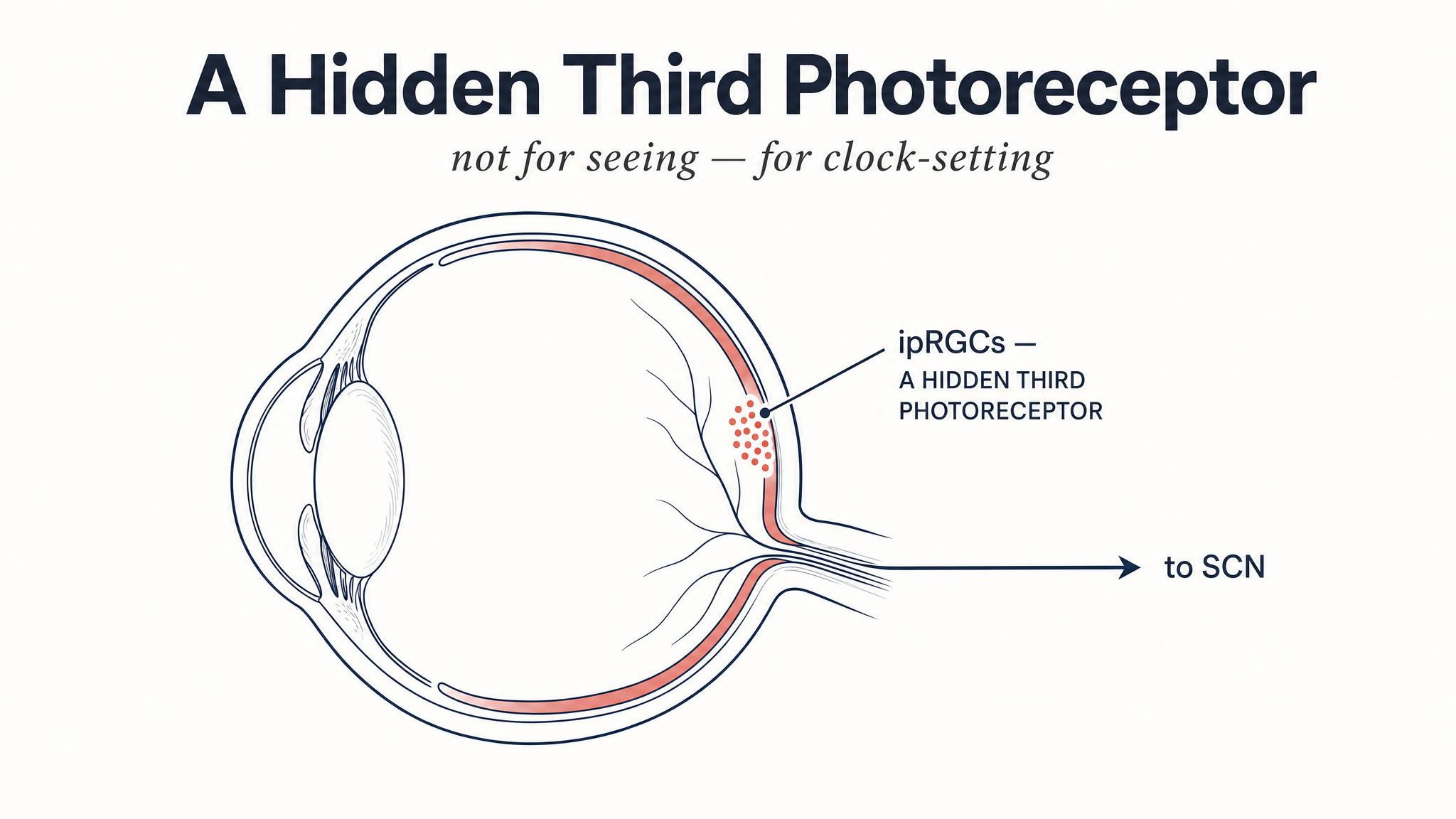
The Hidden Photoreceptors in Your Eye.
Ask most people how many types of light-detecting cells exist in the human eye, and you'll hear two: rods for dim-light vision and cones for color and detail. Until the early two thousands, textbooks agreed. But a series of elegant experiments revealed a third class of photoreceptor hiding in plain sight — intrinsically photosensitive retinal ganglion cells, or I-P-R-G-Cs, as Schmidt, Hattar, and Berson described in 2019.
These cells are rare, making up only about one to two percent of all retinal ganglion cells, and they look nothing like rods or cones. They sit in the ganglion cell layer — the layer closest to the front of the eye — and extend long, branching dendrites that form a wide net across the retina, perfectly designed to catch light from a broad area rather than focusing on fine detail. Most importantly, they contain their own photopigment: MELANOPSIN.
[short pause]
Melanopsin: Not About Seeing, But About Timing.
Melanopsin is an ancient molecule, more closely related to the photopigments of invertebrate eyes than to the rhodopsin in your rods. When photons strike melanopsin, the I-P-R-G-C depolarizes — slowly. While a rod or cone can respond to a flash of light in milliseconds, an I-P-R-G-C takes seconds to fully activate and can remain firing for minutes after the light disappears. As Schmidt, Hattar, and Berson explained in 2019, this sluggishness is a feature, not a bug. The I-P-R-G-C doesn't care about the flicker of a candle or the flash of a passing headlight. It integrates light over long durations, effectively asking a single question: how bright is the world right now?
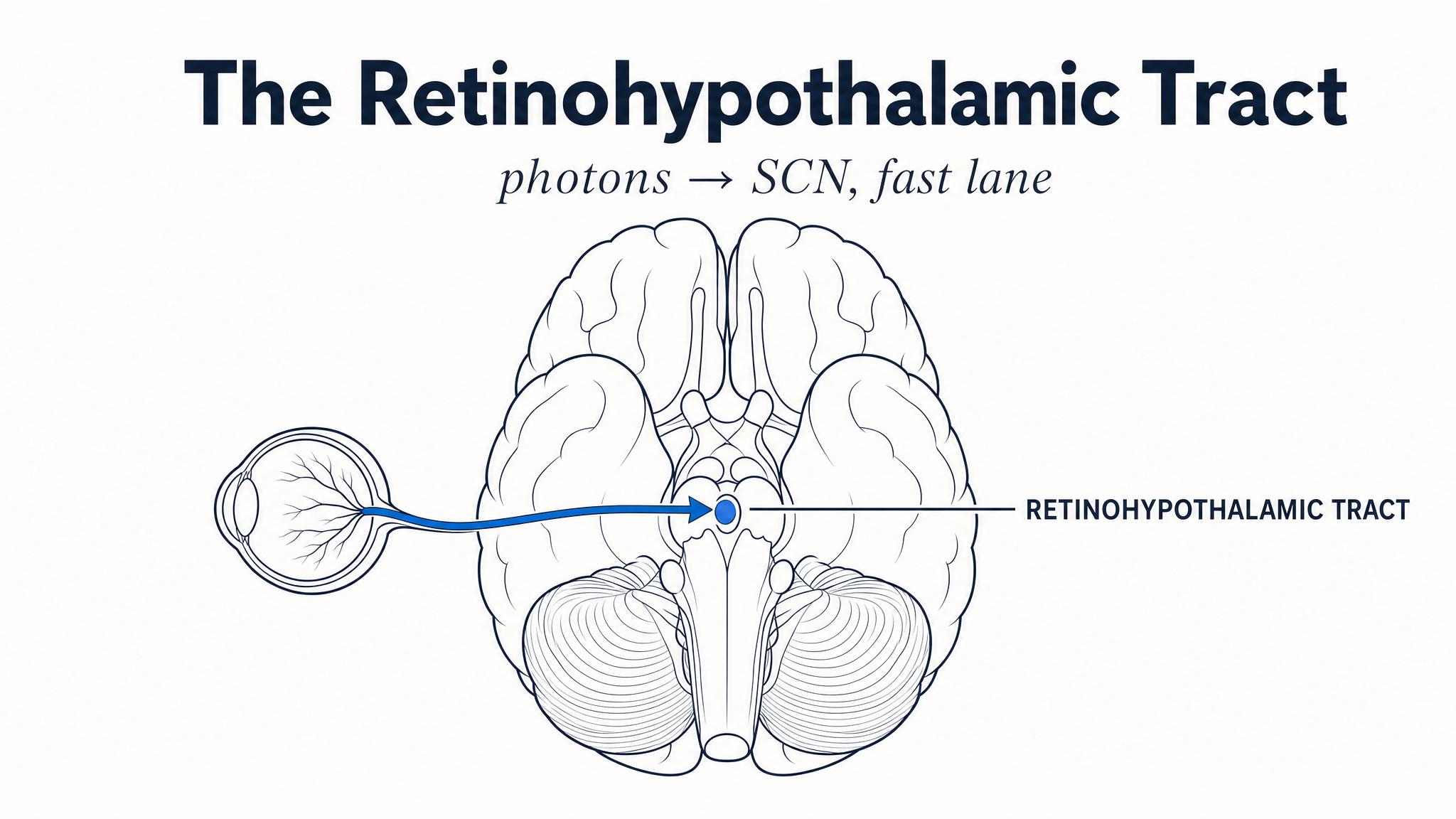
The answer to that question travels along a dedicated neural highway — the retinohypothalamic tract — directly to the suprachiasmatic nucleus, or S-C-N, the master clock you met in Chapter One. This is a monosynaptic connection: one cell, one synapse, one destination. It is among the most direct sensory pathways in the entire brain, and its purpose is simple and singular: to synchronize your internal clock to the external light-dark cycle.
[short pause]
Consider this: People who are completely blind due to damage to their rods and cones can sometimes still entrain their circadian rhythms to the light-dark cycle, while people who lose their eyes entirely cannot. What does this tell you about the role of I-P-R-G-Cs versus rods and cones in circadian photoentrainment? The I-P-R-G-Cs sit in the ganglion cell layer of the retina and project directly to the S-C-N via the retinohypothalamic tract — a dedicated pathway for circadian photoentrainment.
[short pause]
Not All Light Is Equal: Lux, Spectrum, and the Thresholds That Matter.
Understanding that a specialized system detects light for your clock is only half the story. The next question is: how much light does this system need? The answer turns out to be surprisingly specific, and it exposes a critical mismatch between the modern indoor environment and the world our circadian systems evolved to expect.
[short pause]
The Lux Scale: You Probably Aren't Getting Enough.
Light intensity is measured in LUX — the amount of luminous flux per square meter striking a surface. Here is where intuition fails most people. A brightly lit office feels, subjectively, very bright. But in terms of the lux values that matter to your I-P-R-G-Cs, it's barely a whisper. Candlelight provides around ten lux. A typical living room, fifty to one hundred fifty lux. A bright office or classroom, two hundred to five hundred lux. An overcast outdoor day, two thousand to ten thousand lux. Direct sunlight, thirty thousand to one hundred thousand lux.
[short pause]
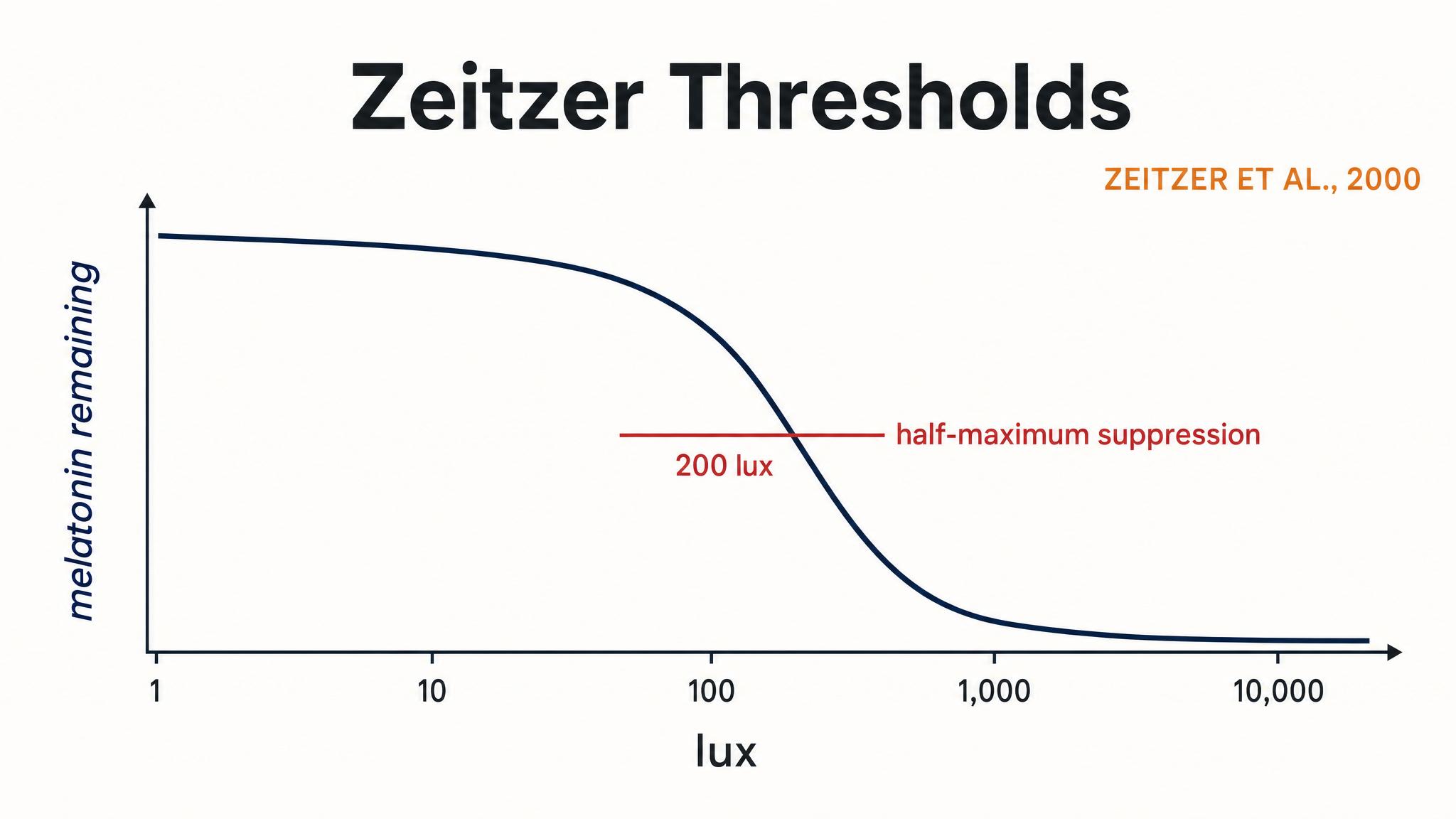
Notice the scale. Even an overcast day delivers ten to fifty times more light than a well-lit office. The landmark study by Zeitzer, Dijk, Kronauer, Brown, and Czeisler in 2000 established that the human circadian system follows a non-linear illuminance-response curve. The half-maximum response for melatonin suppression occurs at roughly one hundred to one hundred twenty lux — meaning ordinary room light can affect the system — but the response saturates around two hundred lux for suppression and approximately five hundred fifty lux for phase shifting. This means that going from one hundred lux, a dim room, to five hundred lux, a bright office, has a much larger circadian impact than going from five hundred lux to five thousand lux. But here's the catch: to get a strong, reliable entrainment signal — the kind that shifts the clock consistently — researchers typically use twenty-five hundred lux or more, and natural daylight effortlessly delivers this.
As Wright and colleagues reported in 2013, their camping participants received roughly thirteen times more daytime light exposure than they did during normal electrical-lighting conditions. This enormous gap is the norm, not the exception. Research by Dunster and colleagues in 2023, tracking over five hundred university students across all four seasons in Seattle, found that each additional hour of daytime light exposure above fifty lux advanced students' chronotype by approximately thirty minutes. Daytime light was a stronger predictor of sleep timing than evening light exposure.
[short pause]
Spectral Sensitivity: The Blue-Light Story Is More Complicated Than You Think.
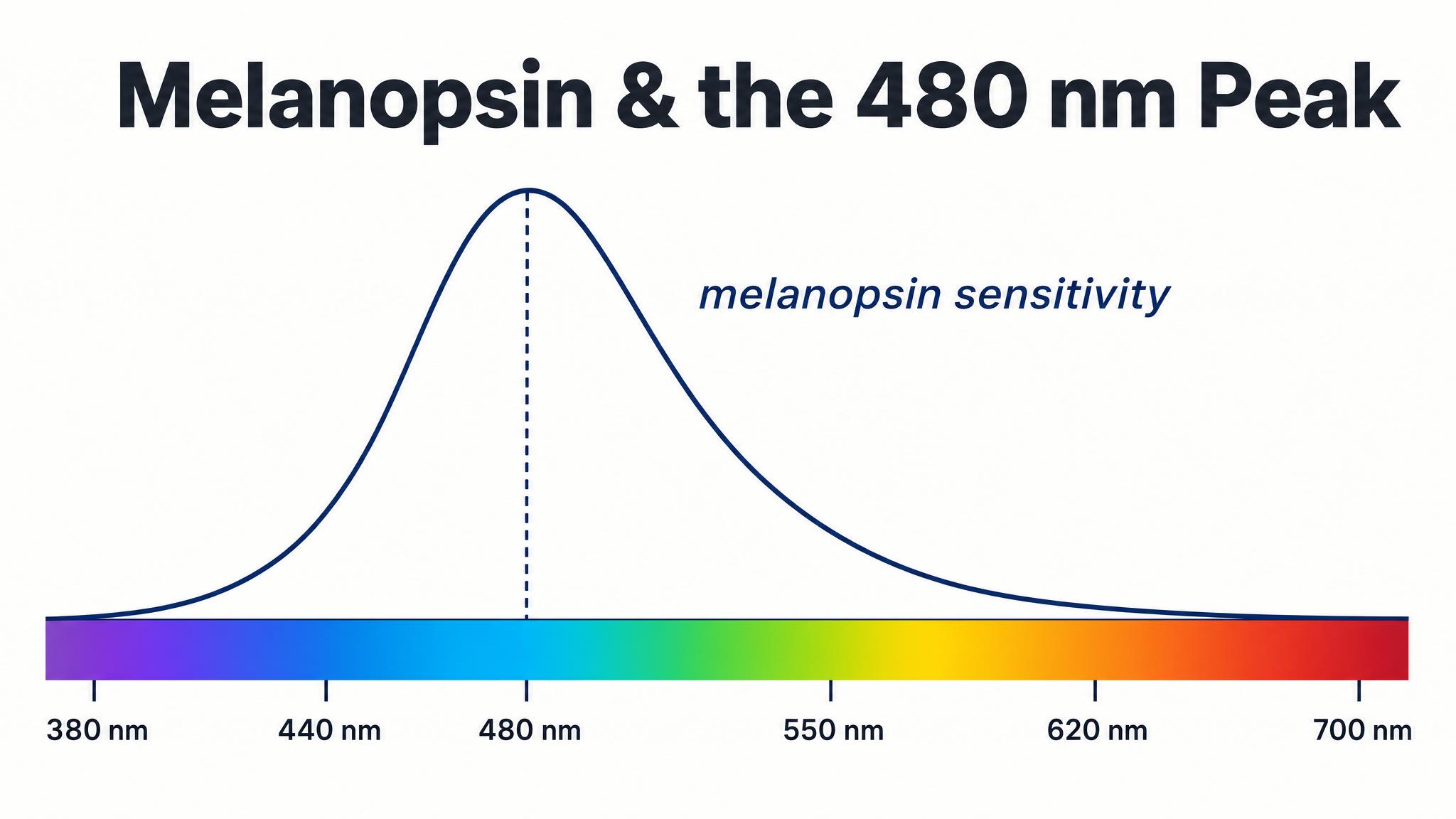
Melanopsin's peak sensitivity falls at approximately four hundred eighty nanometers — squarely in the blue portion of the visible spectrum, as Schmidt, Hattar, and Berson noted in 2019. This is the origin of the now-ubiquitous blue light warnings about screens and L-E-D bulbs. But recent research has complicated this simple narrative considerably.
According to St. Hilaire and colleagues in 2022, who exposed one hundred participants to monochromatic light of different wavelengths, spectral sensitivity changes dynamically over time. During the first minutes of light exposure, the short-wavelength-sensitive S-cones contribute significantly to circadian responses. As exposure continues over hours, melanopsin — with its sluggish, sustained response — dominates. In other words, the system is broadly responsive across wavelengths, especially at higher intensities and longer durations. Blue light isn't magic; bright light is what matters most.
[short pause]
If melanopsin peaks at four hundred eighty nanometers but the circadian system is broadly responsive across wavelengths, why might blue-light-blocking glasses have less impact on circadian entrainment than their marketing suggests? What variable might matter more than the spectral composition of the light? The answer is intensity — the total amount of light matters far more than its color.
[short pause]
The Phase-Response Curve: When You See Light Changes Everything.
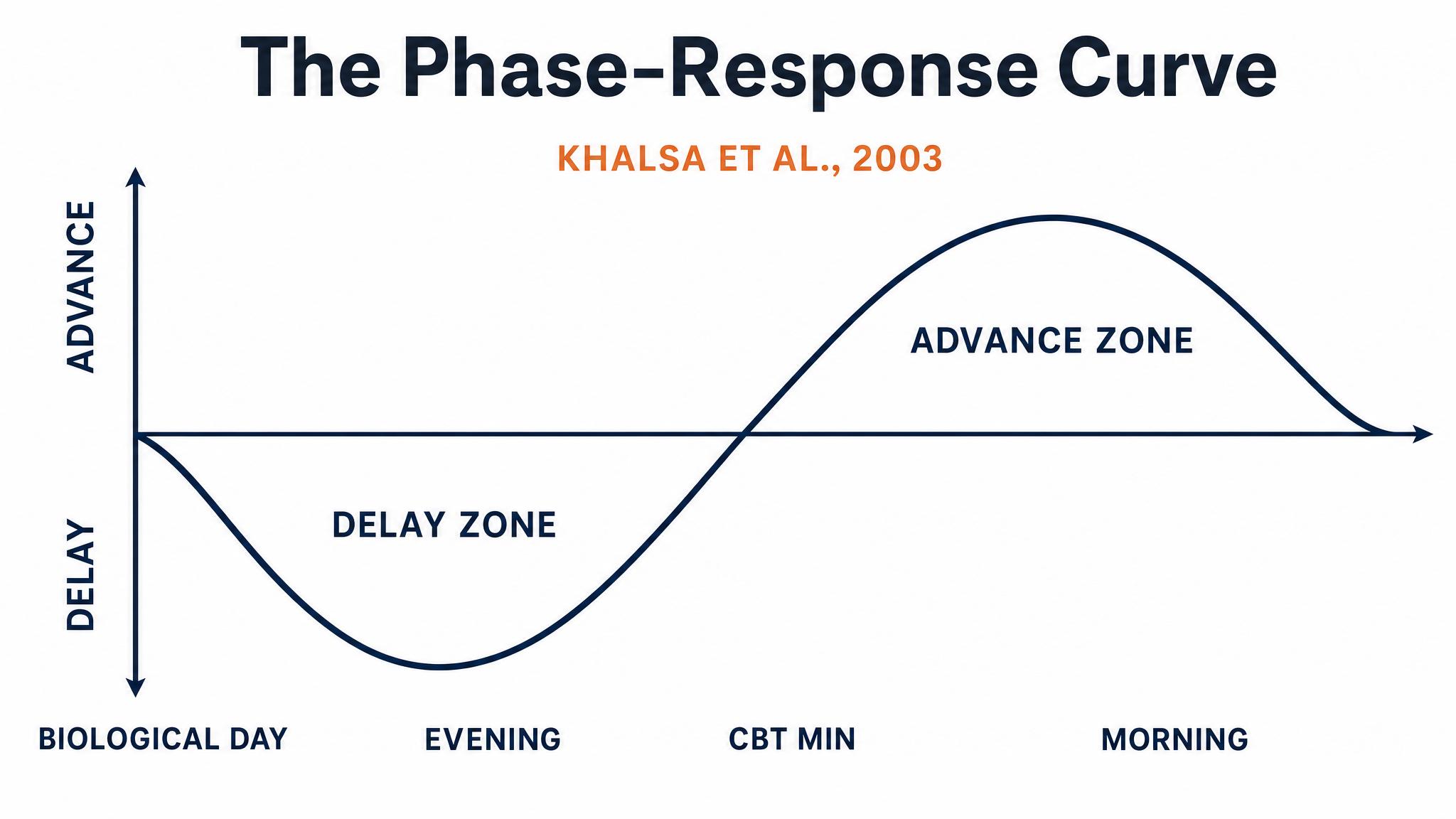
So light is the primary input to your circadian clock, and brighter is generally stronger. But here is the truly consequential insight: the same light stimulus can push your clock in completely opposite directions depending on when it arrives. This relationship is captured in one of the most important graphs in circadian biology: the PHASE-RESPONSE CURVE, or P-R-C.
The definitive human phase-response curve was established by Khalsa, Jewett, Cajochen, and Czeisler in 2003, who exposed twenty-one participants to six point seven hour pulses of bright light at approximately ten thousand lux at different circadian phases. The results were strikingly clear.
[short pause]
Light before the CORE BODY TEMPERATURE MINIMUM, or C-B-T-min, which typically occurs one to two hours before your natural wake time, causes phase delays — your clock shifts later. Light after the C-B-T-min causes phase advances — your clock shifts earlier. The strongest effects occur in the hours immediately surrounding the C-B-T-min, with the total range spanning over five hours of possible shift. Contrary to earlier assumptions, there is no extended dead zone during the subjective day — light at all times has some effect, though it is weakest during the afternoon.
[short pause]
This means that for a typical person waking at seven A-M, with a C-B-T-min around five A-M, bright light at six to eight A-M will advance the clock — making you naturally sleepy and wakeful earlier the next day. But bright light at three to five A-M, say, from a bathroom trip with the lights blazing, will delay the clock, pushing everything later. Same light, opposite effects, separated by just a couple of hours.
The human phase-response curve to bright light shows that light before the core body temperature minimum delays the clock, while light after it advances the clock, based on data from Khalsa and colleagues in 2003.
[short pause]
Morning Light: Your Most Powerful Circadian Tool.
With the phase-response curve in mind, the practical implications become clear. For the vast majority of people — especially those who struggle to wake up in the morning or whose sleep timing has drifted later than desired — bright morning light is the single most effective circadian intervention available.
Crowley and Eastman tested this directly in 2015. They exposed adults to morning bright light at approximately five thousand lux for varying durations combined with afternoon melatonin. Even just thirty minutes of morning bright light produced a one point eight hour phase advance — roughly seventy-five percent of the maximum effect achieved with a full two-hour exposure. The message is encouraging: you don't need to spend hours outdoors. A focused fifteen to thirty minute exposure to bright light shortly after waking can produce meaningful shifts.
[short pause]
The Morning Light Protocol.
A practical morning light strategy involves three elements. First, timing: within the first one to two hours after your natural wake time. This places the light squarely in the phase-advance zone, after your C-B-T-min. Second, intensity: aim for at least two thousand lux. Outdoor light, even on an overcast day, easily exceeds this. If you cannot get outdoors, a ten thousand lux light therapy box positioned twelve to sixteen inches from your face achieves comparable levels. Third, duration: fifteen to thirty minutes is a practical minimum; longer is better but with diminishing returns after about sixty minutes.
The beauty of this approach is its simplicity. Walking to class, eating breakfast near a window, or spending the first twenty minutes of a study session outdoors can all serve as effective light exposure — no specialized equipment required.
[short pause]
Evening Light: Protecting Your Melatonin Onset.
The other side of the phase-response curve tells an equally important story. Light in the hours before and around your C-B-T-min — roughly from the evening through the early night — falls in the phase-delay zone. Bright light during this window pushes your clock later and suppresses the rising tide of melatonin that signals your body to prepare for sleep.
As Zeitzer and colleagues showed in 2000, melatonin suppression saturates at surprisingly low intensities — around two hundred lux. This means that very bright indoor environments in the evening, like well-lit kitchens or fluorescent-lit study spaces, can meaningfully affect your circadian timing. The practical protocol here is straightforward.
[short pause]
First, dim your environment in the two to three hours before your target bedtime. This doesn't require sitting in darkness — fifty lux or below, roughly equivalent to a few dim lamps, is a reasonable target. Second, avoid bright overhead lighting. Overhead lights are particularly effective at reaching the lower retina, where I-P-R-G-C dendrites are dense. Table lamps and floor lamps positioned below eye level deliver less circadian stimulation. Third, prioritize morning light to compensate. As Dunster and colleagues found in 2023, each additional hour of daytime light advanced chronotype by thirty minutes, while each additional hour of post-dusk light delayed it by only fifteen minutes. Strong daytime exposure partially buffers against evening exposure.
[short pause]
A Necessary Nuance About Screens.
This is where we must be honest about the limits of the evidence. Yes, screens emit light with a blue spectral component. Yes, melanopsin is sensitive to blue wavelengths. But a phone held at arm's length typically delivers only thirty to eighty lux to the eye — far below the approximately two hundred lux saturation point for melatonin suppression. The circadian effect of a dim screen is real but modest, especially compared to the effect of overhead room lighting or bright bathroom lights. We will explore the complex relationship between screens and sleep in much greater depth in Chapter Six. For now, the key point is this: the biggest light-related mistake most people make is not that they look at their phones at night — it's that they don't go outside in the morning.
[short pause]
Recall the Wright camping study from 2013: participants received thirteen times more daytime light than in their normal lives. If you only addressed one side of the light equation — morning brightness versus evening dimness — which would likely have the larger impact on your circadian timing, and why? The answer is morning brightness. Daytime light has approximately twice the predictive power for sleep timing as evening light.
[short pause]
Light Meets Chronotype: Personalizing the Signal.
In Chapter One, you identified your chronotype — the genetically influenced tendency to prefer earlier or later sleep timing. Now we can add a crucial layer: chronotype sets the baseline, but light exposure shapes where that baseline actually lands.
Consider two students. One is a natural owl with an M-S-F-s-c, or midpoint of sleep on free days corrected, of five thirty A-M — a late midpoint of sleep. The other is a lark with an M-S-F-s-c of two thirty A-M. The phase-response curve operates identically in both, but because their C-B-T-min occurs at different clock times, the same environmental light hits them at different circadian phases. For the owl whose C-B-T-min might be at eight A-M, light at seven A-M actually falls in the delay zone — making things worse. This same owl needs light after eight A-M to get an advance signal, which is exactly when they'd naturally be waking up if their schedule allowed it.
[short pause]
This reframes the Dunster Seattle school start-time study from Chapter One, conducted in 2018. When school start times shifted from seven fifty to eight forty-five A-M, students gained thirty-four minutes of sleep — but the mechanism may be more nuanced than simply more time in bed. The later start allowed many students, particularly owls, to receive their first bright light exposure at a circadian phase that fell after their C-B-T-min rather than before it. The light they were getting shifted from the delay zone to the advance zone — amplifying rather than fighting their biology. Morning light exposure patterns did shift in the study, as Dunster and colleagues reported in 2018, consistent with this interpretation.
[short pause]
The same morning light hits larks and owls at different circadian phases. An owl forced to wake early may receive light in the delay zone, inadvertently pushing their clock even later. The practical lesson is that a one-size-fits-all morning light prescription misses this individual variation. If you're an extreme owl, your optimal light-exposure window is later in the morning — but you may need to start with light at whatever time you do wake up and gradually shift earlier as your clock moves. Think of it as meeting your biology where it is, then using light to nudge it in the direction you want.
[short pause]
Putting It All Together: Light as a Lifestyle Variable.
The core message of this chapter can be distilled into a simple hierarchy. The most impactful light-related change most people can make is getting more bright light during the day, particularly in the morning. The second most impactful change is dimming the environment in the evening. The third — and often the one that gets the most attention — is managing screen light, which matters but much less than the first two.
This hierarchy aligns beautifully with the data. As Dunster and colleagues found in 2023, daytime light was twice as powerful as evening light in predicting sleep timing among university students. Wright and colleagues demonstrated in 2013 that natural light alone could compress chronotype differences and advance the clock by two hours in a week. And Crowley and Eastman showed in 2015 that even thirty minutes of bright morning light can produce clinically meaningful phase advances.
[short pause]
The most effective sleep intervention available to most people costs nothing, requires no prescription, and takes less than thirty minutes: step outside in the morning.
[short pause]
Of course, the real world isn't a research protocol. Winters at high latitudes mean sparse morning light. Shift workers face impossible schedules. Students cramming in windowless libraries miss daylight entirely. These complications are real, and we'll return to them in later chapters. But the underlying biology is now clear: your circadian clock runs on light, and the modern indoor lifestyle starves it of the signal it needs most.
[short pause]
Key Takeaways.
A specialized class of retinal cells — I-P-R-G-Cs containing melanopsin — detects ambient light levels and signals the S-C-N directly, independent of conscious vision.
Melanopsin peaks at approximately four hundred eighty nanometers, in the blue range, but is broadly responsive; bright light of any color is more important than spectral composition alone.
Indoor environments at one hundred to five hundred lux are dramatically dimmer than outdoor daylight at two thousand to one hundred thousand lux, creating a chronic circadian signal deficit for people who spend most of their time indoors.
The phase-response curve determines whether light advances or delays your clock: light after the core body temperature minimum, roughly after natural wake time, advances the clock; light before it delays the clock.
Morning bright light for fifteen to thirty plus minutes at two thousand lux or more is the single most effective tool for advancing the circadian clock and stabilizing sleep timing.
Evening light dimming protects melatonin onset, but daytime light has approximately twice the predictive power for sleep timing as evening light.
The same environmental light hits different chronotypes at different circadian phases — personalized light timing matters.
Screen light at arm's length, thirty to eighty lux, has a real but modest circadian effect compared to room lighting and especially outdoor light.
[short pause]
Looking Ahead.
You now understand the most powerful external input to your circadian clock. In Chapter Four, we'll turn to the internal partner: sleep pressure and the homeostatic drive. While light sets the timing of sleep, adenosine accumulation sets its urgency. Together, these two forces — the circadian rhythm and the sleep homeostat — form the two-process model that explains not just when you sleep, but why you sometimes can't stay awake. We'll also explore the molecule that blocks adenosine's signal: caffeine — and why its timing matters more than its dose.









The Caffeine Equation — The World's Most Popular Sleep Saboteur
With circadian biology, sleep architecture, and light exposure established, the course now introduces the molecule that most directly interferes with Process S: caffeine. This chapter reveals caffeine's mechanism of…
Is it ever actually worth pulling an all-nighter with coffee, or are you just making tomorrow worse for today's panic?
Hot take
What's your hard cutoff time for caffeine? 2pm? 4pm? 'I drink espresso at midnight and sleep fine'?
Quick poll
Would you rather: Give up coffee/energy drinks completely OR give up one hour of sleep every night forever?
Would you rather
We all have that friend who chugs Red Bull at 10pm and passes out by 11. Are they superhuman or are they lying?
Real talk
You have a huge exam at 8am. Do you: Stay up late cramming with coffee, or go to bed early and risk not covering everything?
What would you do?
Coffee before a nap — genius life hack or absolute waste of coffee?
Debate time
Your Caffeine Half-Life Timeline
12-15 minutesStudents document their caffeine intake from the past 24 hours (time + amount). They calculate remaining caffeine in their system using half-life rules (5-6 hours). Then turn to neighbors (2-3 people) and compare: Who would sleep better tonight? Who's still 50%+ caffeinated at bedtime? Groups report out surprising findings. Instructor highlights the metabolizer variation: 'Some of you clear this in 3 hours, others take 9 hours—same coffee, different genes.'
Adenosine Accumulation Auction
10-12 minutesInstructor presents 5 scenarios on screen (e.g., '8am coffee + 3pm coffee', '2pm energy drink', 'no caffeine today', '10pm tea'). Students 'bid' on which scenario will have the MOST adenosine build-up by 11pm by standing/raising hands. After each vote, instructor reveals the answer and explains receptor antagonism dynamics: caffeine doesn't stop adenosine accumulation, it just blocks the receptors—so when caffeine wears off, you get hit with the backlog. Discuss why late-day caffeine is particularly problematic.
Fast vs. Slow Metabolizer Debate
15-18 minutesDivide room by rows: left half are 'Fast Metabolizers' (efficient CYP1A2), right half are 'Slow Metabolizers' (poor CYP1A2). Present 3 caffeine strategies on screen (e.g., 'afternoon coffee at 2pm', 'double espresso at noon only', 'small amounts throughout day'). Each side discusses with neighbors for 2 minutes, then representatives from each side argue FOR or AGAINST each strategy from their genetic perspective. Class votes on which strategy wins for each metabolism type. Reveal real research on athletic performance and sleep outcomes for each genotype.
Deep Sleep Detective: N3 Stage Analysis
12-15 minutesDisplay two sleep hypnograms on screen: Subject A (no caffeine) and Subject B (200mg caffeine at 5pm). Students work with neighbors to identify differences, specifically in N3 deep sleep duration and timing. After 4-5 minutes, collect observations from around room. Reveal the key finding: caffeine selectively impacts N3 in the first sleep cycle, even when total sleep time is similar. Discuss why this matters—N3 is critical for physical restoration, memory consolidation, immune function. Students then estimate: if Subject B had stopped caffeine at 2pm instead, would N3 recover? Calculate together using half-life.
The Caffeine Optimization Challenge
10-12 minutesPresent 3 fictional students with different schedules and goals: Student A (athlete needing performance boost and recovery), Student B (exam tomorrow, needs alertness but good sleep), Student C (night shift worker). Students work with neighbors to design optimal caffeine timing and dosing for each person. Write recommendations on paper/phone. Instructor collects 4-5 responses verbally from different sections, then reveals 'expert recommendations' based on research. Highlight trade-offs: adenosine buildup, half-life timing, N3 protection, individual variation.
The 11pm Caffeine Census
8-10 minutesReal-time class data collection: students stand up if they've had caffeine after 4pm today (or yesterday if morning class). Those standing share with neighbors what they consumed and when. Instructor does live calculation on board: '30 students standing, average 200mg at 5pm, half-life of 5-6 hours... that means at 11pm, 20-25 of you still have 50-100mg in your system—equivalent to half a cup of coffee AT BEDTIME.' Visual representation: 'If I handed you coffee right before bed, you'd refuse—but functionally, that's what you did.' Students predict their own sleep quality tonight based on their data. Follow up next class: 'How did you actually sleep?'
Transcript
It's two thirty in the afternoon, and you're fading. Your eyelids are heavy, your focus is crumbling, and the afternoon stretches ahead like an endless hallway. So you do what roughly eighty-five percent of adults worldwide do: you reach for caffeine. Maybe it's a latte, maybe it's a tea, maybe it's an energy drink. Within thirty minutes, the fog lifts. You feel sharp again, productive, even energized. The problem seems solved.
That evening, you climb into bed at eleven P M. You fall asleep without too much trouble. You wake after seven hours. And yet somehow, you feel unrested, groggy, heavy, as if sleep didn't quite do its job. So you reach for that morning coffee a little more eagerly, and by mid-afternoon, the cycle begins again. What's happening behind the scenes is a story of molecular deception: caffeine didn't eliminate your tiredness that afternoon, it hid it. And when you slept, it quietly stole the deepest, most restorative stage of your night. This chapter unpacks how.
[short pause]
From Chapters one through three to your coffee cup. Over the first three chapters, you've built a sophisticated model of sleep regulation. In Chapter one, you learned about Process S, the homeostatic sleep drive that rises the longer you stay awake, like a pressure gauge slowly filling. In Chapter two, you explored sleep architecture, the structured progression through N-one, N-two, N-three, and REM stages, each serving distinct biological functions. In Chapter three, you saw how light sculpts your circadian rhythm, the internal clock that tells your body when to sleep. Now we introduce the single most common chemical that disrupts this elegant system: caffeine.
Caffeine is not merely a stimulant in the colloquial sense. It is, with striking molecular precision, an ADENOSINE RECEPTOR ANTAGONIST, a chemical that targets the exact signaling pathway that Process S relies upon. To understand why your afternoon coffee can silently undermine tonight's sleep, we need to return to the molecule at the heart of sleep pressure: adenosine.
[short pause]
The Adenosine Story: How Your Brain Builds Sleep Pressure. Recall from Chapter one that adenosine is a byproduct of neuronal energy metabolism. Every time your brain cells fire, processing information, solving problems, navigating your day, they consume adenosine triphosphate, or A-T-P, for fuel, and adenosine gradually accumulates in the extracellular space. This accumulation is the biological basis of Process S. The longer you're awake, the more adenosine builds up, and the sleepier you feel, as Reichert and colleagues found in 2022.
Adenosine produces this sensation of drowsiness by binding to specific receptors on the surface of neurons, primarily the A-one receptors, which inhibit wake-promoting neurons, and the A-two-A receptors, which activate sleep-promoting pathways in the basal forebrain. When adenosine docks into these receptors, it's like turning a dimmer switch. Neural activity gradually winds down, arousal decreases, and the brain moves toward a sleep-ready state.
Here is where caffeine enters the picture, and where its mechanism is both elegant and insidious.
[short pause]
A Molecular Game of Musical Chairs. Caffeine's three-dimensional molecular structure is remarkably similar to adenosine. Similar enough, in fact, that caffeine can slide into the same A-one and A-two-A receptors. But here's the critical difference: caffeine occupies those receptors without activating them. It sits in the chair but doesn't do the job. This is what pharmacologists mean by ANTAGONISM: caffeine blocks the receptor without triggering the downstream signal, as Reichert and colleagues described in 2022.
Imagine a game of musical chairs. Adenosine molecules are the players trying to sit down, and every time one sits, the sleepiness signal gets a little louder. Caffeine molecules are imposters wearing adenosine costumes. They rush in and occupy chairs, preventing real adenosine from sitting. The music keeps playing, the signal stays quiet, and you feel awake. But, and this is the point students often miss, adenosine production doesn't stop. Your brain is still consuming energy, still generating adenosine. The molecules are still accumulating. They just can't get to their chairs.
This is why caffeine doesn't give you energy in any true metabolic sense. It gives you the perception of wakefulness by masking a signal that was telling you the truth: you're tired, and getting more tired by the minute.
[short pause]
Consider this: if caffeine blocks adenosine receptors without reducing adenosine accumulation, what do you predict happens when caffeine is eventually metabolized and clears those receptors? Think back to the Process S curve from Chapter one. What would the curve look like if the signal were suddenly unmasked?
[short pause]
The Caffeine Crash, Explained. You already know the answer from experience. When caffeine is eventually metabolized by the liver and its molecules release from the receptors, all of that pent-up adenosine, which has been accumulating for hours, suddenly floods the now-vacant receptors. The dimmer switch doesn't gently descend; it slams down. This is the caffeine crash: not a new wave of tiredness, but the delayed arrival of tiredness that was building all along. The Process S curve didn't flatten while you were caffeinated, it kept rising in the background, invisible. Caffeine merely shifted when you felt it, as Reichert and colleagues explained in 2022.
[short pause]
Caffeine Pharmacokinetics: The HALF-LIFE Problem. Understanding that caffeine masks adenosine is only half the story. The other half is understanding how long it takes your body to clear caffeine, because this determines whether that afternoon coffee is still lingering in your brain at bedtime.
Caffeine is absorbed rapidly through the gastrointestinal tract, reaching peak blood concentration within thirty to sixty minutes of ingestion. It readily crosses the blood-brain barrier, meaning it reaches your central nervous system quickly and efficiently. But clearing it is a far slower process. The half-life of caffeine, the time it takes for your body to eliminate half of the caffeine in your bloodstream, averages approximately five hours in healthy adults, though the range spans from one and a half to nine and a half hours, according to the Institute of Medicine in 2001.
Let's do some practical arithmetic. Say you drink a standard drip coffee containing two hundred milligrams of caffeine at two P M. With a five-hour half-life: by seven P M, one hundred milligrams is still circulating, half the original dose. By midnight, approximately fifty milligrams remains, one quarter. By five A M, approximately twenty-five milligrams is still present, one eighth. That single afternoon coffee has measurable caffeine levels in your brain well into the early morning hours. And if you had two coffees, or added an energy drink, the numbers stack up. Caffeine from multiple doses accumulates, and each dose decays on its own timeline.
[short pause]
Why Your Half-Life Isn't Average. The five-hour average conceals enormous individual variation, and this variation is largely determined by the C-Y-P-one-A-two enzyme in your liver, the enzyme responsible for metabolizing approximately ninety-five percent of ingested caffeine, as Nehlig reported in 2018. Your C-Y-P-one-A-two gene comes in different variants, and these variants produce dramatically different metabolic speeds.
Fast metabolizers carry gene variants that produce highly active C-Y-P-one-A-two enzymes. Their half-life may be as short as two to three hours. Slow metabolizers carry less active variants and may have half-lives of seven to nine hours, meaning caffeine lingers in their system nearly three times longer. In genetic knockout studies, mice lacking C-Y-P-one-A-two entirely showed caffeine half-lives seven times longer than normal, confirming this enzyme as the critical bottleneck, as Berthou and colleagues found in 1996.
But genetics isn't the only factor. Several conditions and substances alter caffeine metabolism significantly, according to the Institute of Medicine in 2001 and Nehlig in 2018. Oral contraceptives can approximately double caffeine half-life. Pregnancy extends half-life dramatically, up to fifteen hours in the third trimester. Smoking accelerates C-Y-P-one-A-two activity, shortening half-life, which is why smokers often drink more coffee. Age: older adults generally metabolize caffeine more slowly. And liver disease can extend half-life substantially.
This variation explains a frustrating reality: generic advice like "no coffee after two P M" is reasonable for average metabolizers but may be too permissive for slow metabolizers and unnecessarily restrictive for fast ones. The right answer depends on your biology.
[short pause]
The Deep Sleep Tax: What Caffeine Really Steals. Here is where the caffeine story takes its most consequential turn, and where many people's confidence that "coffee doesn't affect my sleep" starts to crumble.
In a landmark study, Drake and colleagues in 2013 gave participants four hundred milligrams of caffeine at three different times: immediately before bed, three hours before bed, and six hours before bed. The results were striking: even caffeine consumed six hours before bedtime reduced total objective sleep time by more than one hour. But the truly remarkable finding was this: participants themselves did not perceive the disruption. Their subjective reports of sleep quality remained largely unchanged even as their polysomnography readings told a very different story.
[short pause]
Recall from Chapter two that different sleep stages serve different functions: N-three for physical restoration and memory consolidation, REM for emotional processing. If caffeine selectively reduces one stage while preserving total sleep duration, a person could sleep enough hours and still suffer specific functional deficits.
[short pause]
This disconnect between subjective experience and objective measurement is one of the most important takeaways from this chapter. And it becomes even more concerning when we examine which part of sleep caffeine targets.
Caffeine's Selective Attack on N-three SLOW-WAVE SLEEP. A comprehensive meta-analysis by Gardiner and colleagues in 2023 synthesized data across multiple studies and found that caffeine produces significant dose-dependent reductions in N-three, or slow-wave, sleep, an average reduction of eleven point four minutes in absolute N-three time. Four studies additionally found significant reductions in delta-frequency E-E-G power during N-REM sleep, confirming that it's not just time in N-three that decreases but the depth and intensity of slow-wave activity.
This matters enormously because, as you learned in Chapter two, N-three is the stage most associated with physical restoration, growth hormone release, immune function, and the glymphatic clearance of metabolic waste, including beta-amyloid, a protein implicated in Alzheimer's disease. N-three is concentrated in the first two sleep cycles, which is precisely the period when residual caffeine levels are highest if consumed in the afternoon or evening.
Perhaps the most striking demonstration comes from the classic study by Landolt and colleagues in 1995, who showed that caffeine administered at seven ten in the morning, roughly sixteen hours before sleep onset, still significantly reduced sleep efficiency and delta-frequency power during nighttime sleep. Even at a dose of just two hundred milligrams, roughly two cups of coffee, and even when salivary caffeine levels had declined to very low concentrations by bedtime, the residual impact on slow-wave sleep architecture was measurable.
A more recent randomized clinical trial reinforced these findings. Gardiner and colleagues in 2025 demonstrated that four hundred milligrams of caffeine consumed within twelve hours of bedtime significantly reduced N-three sleep, noting "the potent ability of caffeine to reduce N-three sleep." Critically, they found that while our brains normally protect N-three sleep even during sleep restriction, treating it as the highest priority stage, caffeine overrides this protective mechanism. The one reassuring finding: a one hundred milligram dose, roughly one cup of tea or a small coffee, did not produce significant N-three disruption.
[short pause]
"But Coffee Doesn't Affect My Sleep." This is the most common objection students, and adults generally, raise when presented with caffeine-sleep research. And on the surface, it seems like a reasonable claim. Many people drink coffee late in the day, fall asleep without difficulty, and wake up after seven or eight hours.
The study by Drake and colleagues in 2013 directly addresses this. Participants who consumed four hundred milligrams of caffeine six hours before bed reported subjective sleep quality that was essentially unchanged from placebo nights, yet polysomnography revealed they lost more than an hour of total sleep and experienced significantly altered sleep architecture. The participants were, in a very real sense, unaware of the damage.
As Drake and colleagues stated in 2013: "Subjective reports of sleep quality are unreliable indicators of caffeine-induced sleep disruption. Individuals consistently underestimate the effects of caffeine on their sleep when compared to objective polysomnographic measures."
[short pause]
There are several reasons for this perceptual blindness. First, sleep onset may be unaffected. Caffeine's primary impact is on sleep architecture, not necessarily sleep latency. You can fall asleep on schedule and still lose deep sleep. Second, we don't perceive sleep stages. You cannot consciously tell the difference between a night of robust N-three sleep and one where N-three was curtailed by thirty percent. The experience of sleeping feels similar either way. Third, chronic caffeine use resets your baseline. If you drink caffeine daily, you may have never experienced your true sleep architecture as an adult. What feels normal may already be a caffeine-degraded version of your potential sleep quality. And fourth, tolerance masks some wakefulness effects but does not eliminate the sleep architecture disruption. Regular caffeine users develop tolerance to the subjective alerting effects, but research suggests that the N-three-suppressing effects persist, as Reichert and colleagues noted in 2022.
The implication is both humbling and empowering. Humbling because it means our intuitions about our own sleep are less reliable than we'd like. Empowering because it suggests that for many people, a simple adjustment to caffeine timing could unlock meaningfully better sleep, without sleeping more hours, just sleeping more deeply.
[short pause]
Building Your Personal Caffeine Cutoff. Armed with the pharmacology, the individual variation data, and the sleep architecture research, you can now approach caffeine with a degree of precision that most people never apply. The goal isn't necessarily to eliminate caffeine. It's to align your consumption with your biology so that caffeine serves you during the day without sabotaging you at night.
The Practical Formula: a reasonable heuristic is to ensure that your blood caffeine level is below approximately one hundred milligrams equivalent by bedtime. Gardiner and colleagues in 2025 found that one hundred milligram doses did not produce significant N-three disruption, while four hundred milligram doses consumed within twelve hours did. Working backward from your bedtime using your estimated half-life: fast metabolizers, with roughly three-hour half-life, a two hundred milligram coffee at four P M drops to around fifty milligrams by ten P M, likely acceptable for an eleven P M bedtime. Average metabolizers, with roughly five-hour half-life, the same two hundred milligram coffee at four P M is still around eighty milligrams at ten P M and around fifty milligrams at midnight. A noon cutoff for larger doses is safest. Slow metabolizers, with roughly seven and a half hour half-life, two hundred milligrams at noon is still around one hundred milligrams at seven thirty P M and around fifty milligrams at three A M. A morning-only policy may be necessary.
If you don't know your metabolizer type, several clues can help. Do you feel wired for hours after a single coffee? You may be a slow metabolizer. Can you drink espresso at dinner and fall asleep easily? You may be fast, though remember, falling asleep easily doesn't mean your deep sleep is unaffected. Are you taking oral contraceptives, are you over sixty, or do you have liver concerns? Factor in a longer half-life.
[short pause]
Consider your own caffeine habits. At what time do you consume your last caffeinated drink? Using the half-life estimates we've discussed, roughly how much caffeine is still circulating at your typical bedtime? You might be surprised by the amount.
[short pause]
Connecting Back to the Two-Process Model. The beauty of understanding caffeine through the lens of Process S is that it demystifies both its appeal and its risks. Caffeine doesn't give you extra wakefulness, it borrows it from your homeostatic sleep drive, then charges interest in the form of degraded deep sleep. Process S rises linearly during wakefulness. Caffeine doesn't change that line, it merely hides it from your conscious experience while it keeps climbing. The crash, when it comes, is the moment the mask slips and the true height of the curve becomes apparent.
Meanwhile, caffeine also interacts with Process C, the circadian system. Caffeine consumed in the evening can delay the timing of melatonin release, much like the evening light exposure discussed in Chapter three, further pushing your circadian clock later and making it harder to fall asleep at your intended time. The two-process model thus becomes a framework for understanding not just why caffeine keeps you awake, but why it fragments your sleep even when it doesn't prevent it.
[short pause]
Key Takeaways. Caffeine is an adenosine receptor antagonist. It blocks the A-one and A-two-A receptors without activating them, masking sleepiness without reducing the underlying sleep pressure. Adenosine continues to accumulate during caffeine use; when caffeine is metabolized, the pent-up adenosine floods receptors, causing the caffeine crash. Caffeine's half-life averages around five hours but ranges from one and a half to nine and a half hours, driven primarily by C-Y-P-one-A-two genetics, with oral contraceptives, pregnancy, age, and liver health as additional factors. Caffeine disproportionately reduces N-three, slow-wave deep sleep, even when total sleep duration appears normal, by an average of around eleven minutes, with greater effects at higher doses and later timing. Subjective sleep quality is a poor indicator of caffeine's impact; people consistently underestimate the disruption measured by polysomnography. Personalized caffeine cutoffs should account for individual metabolism, total daily intake, and the goal of reaching approximately one hundred milligrams or less circulating caffeine by bedtime. And caffeine tolerance reduces the perceived alerting effects but does not eliminate sleep architecture disruption.
[short pause]
Looking Ahead. Now that you understand how caffeine masks the homeostatic sleep signal, Chapter five will explore what happens when we try to manipulate sleep from the other direction, with alcohol and sedatives. You'll discover that alcohol, like caffeine, deceives us about our sleep quality, but through a completely different mechanism: it sedates the brain in the first half of the night and then fragments sleep architecture in the second half. The question of "why does a nightcap make me feel sleepy but leave me exhausted?" connects directly back to the architecture you've studied and the Process S dynamics caffeine has now revealed.

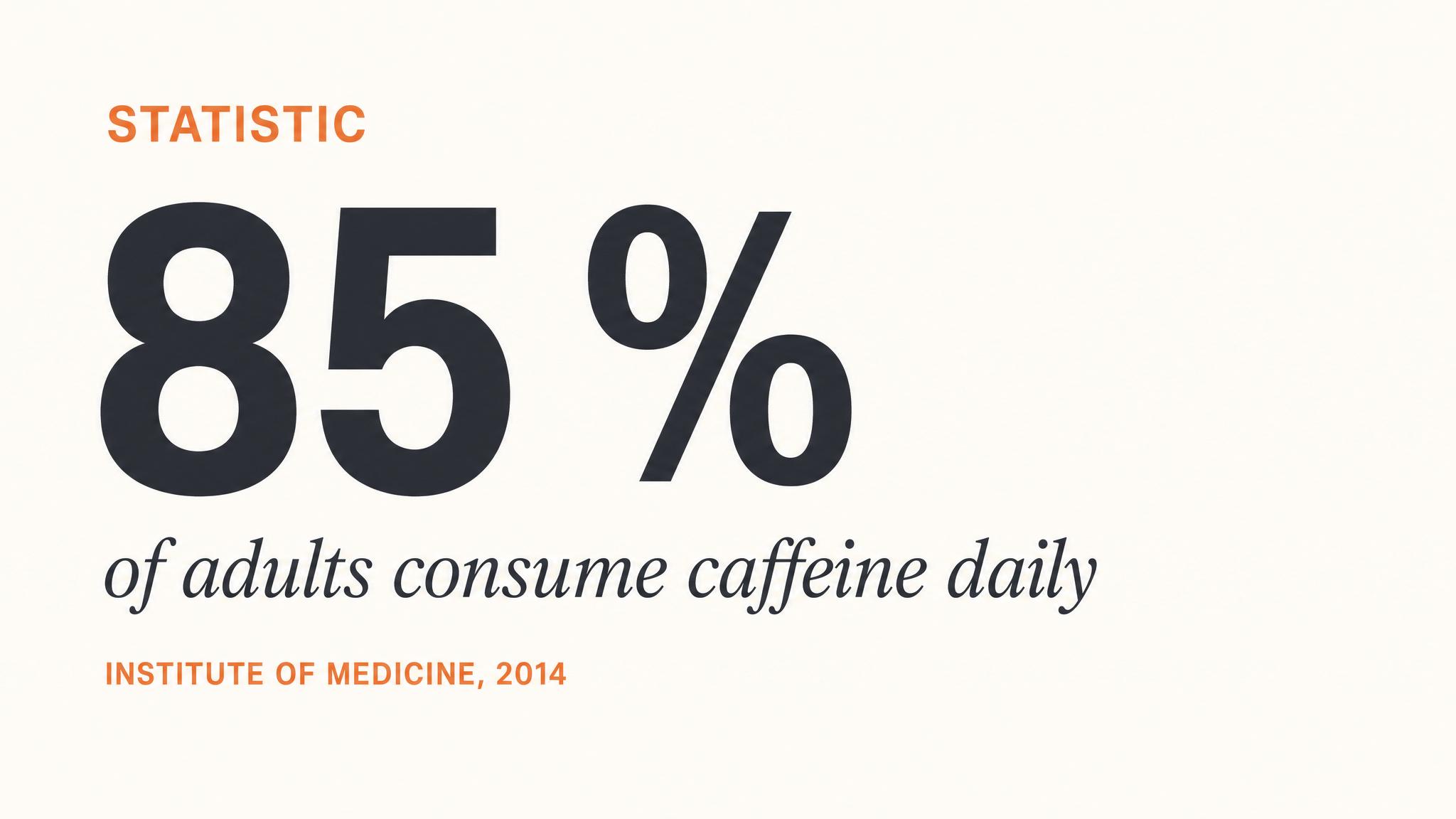
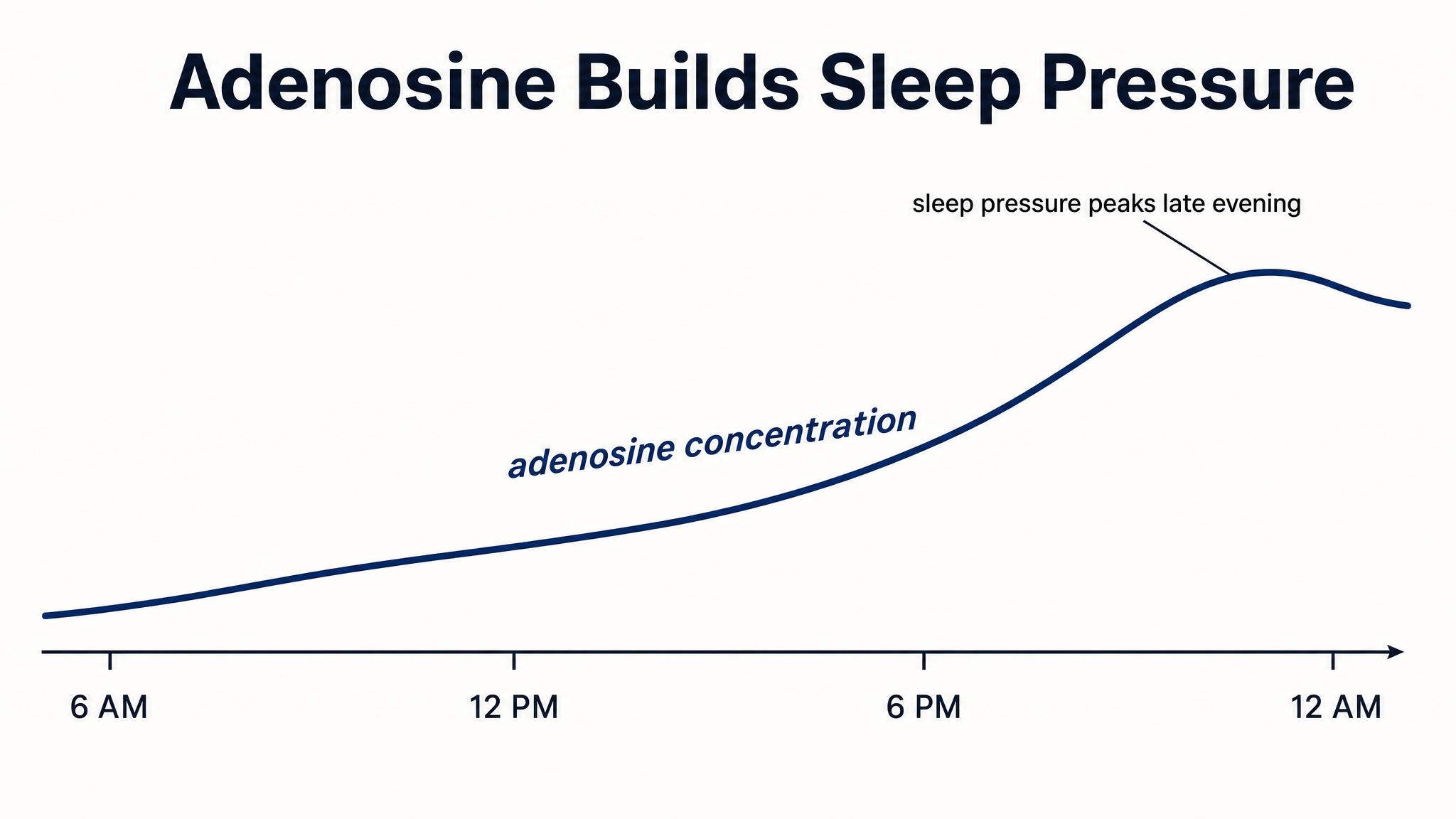
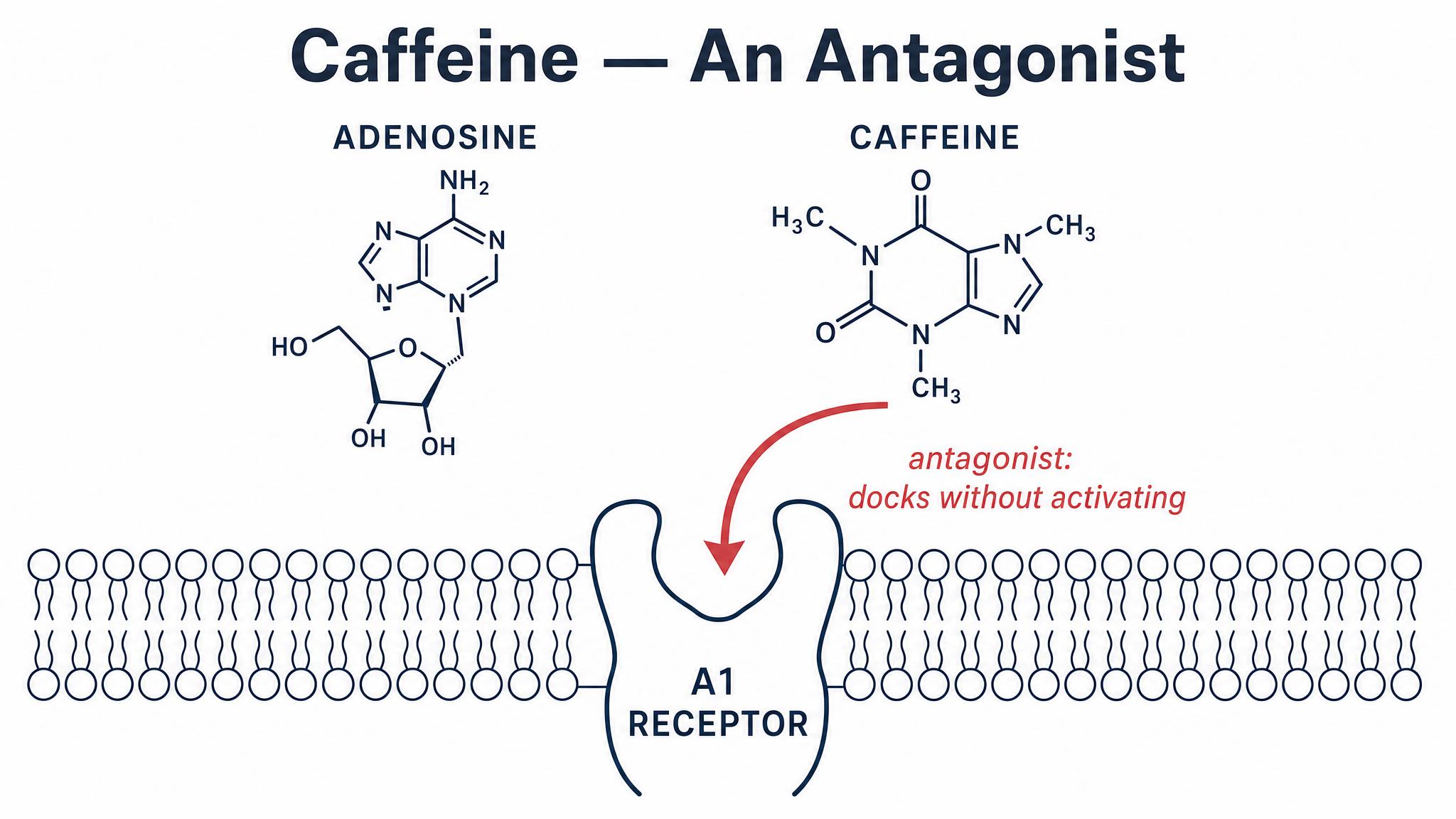

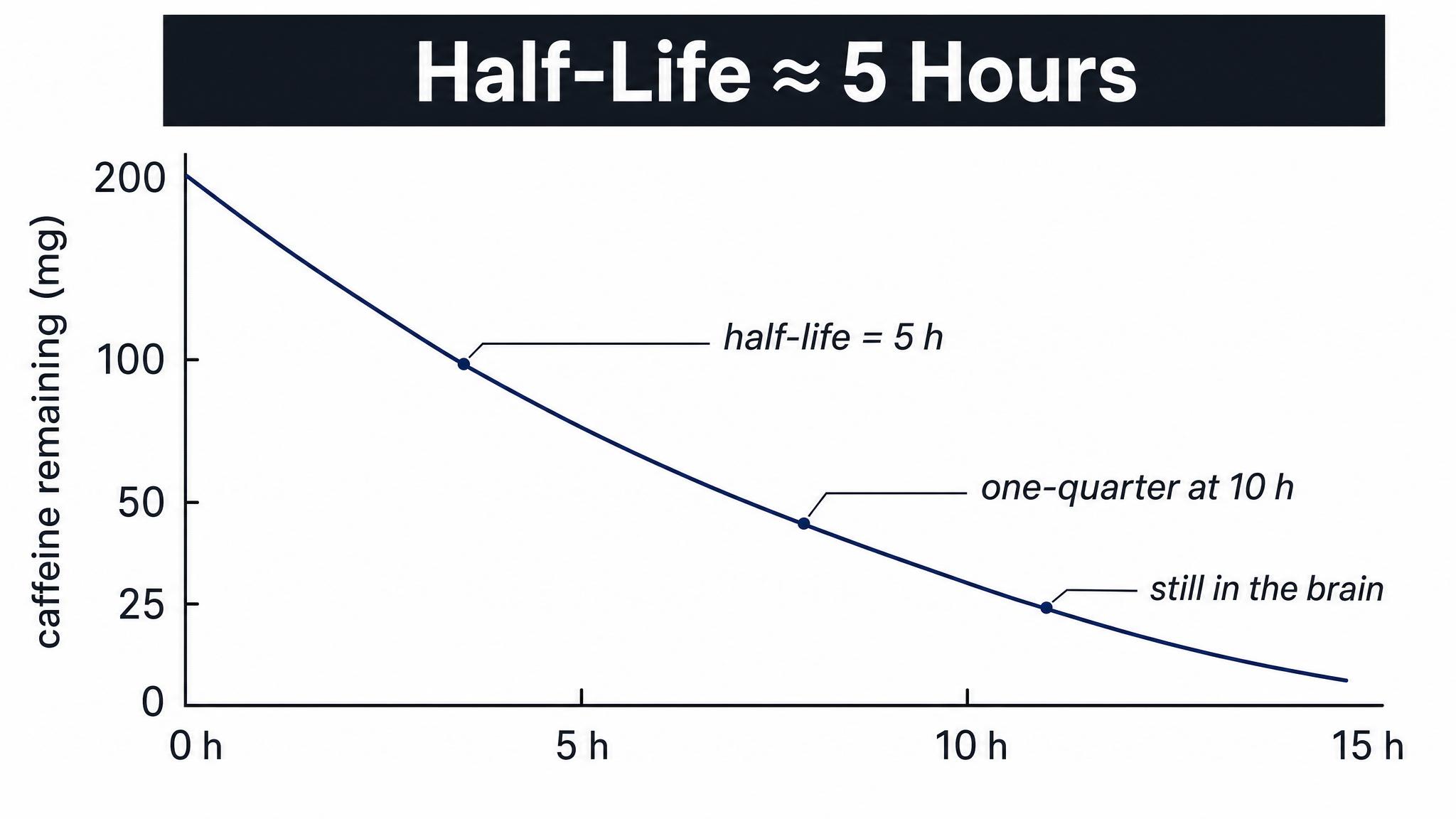

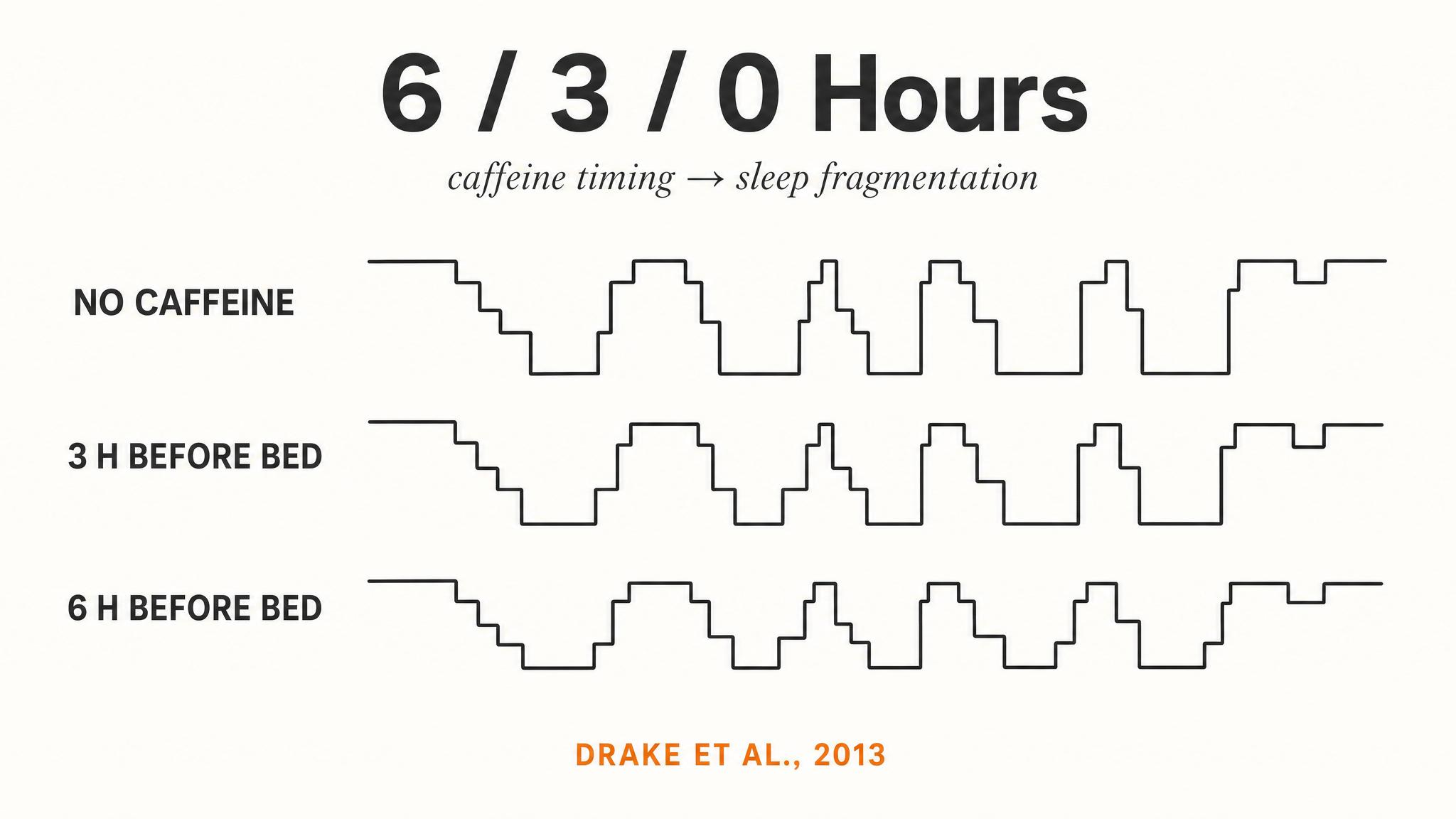

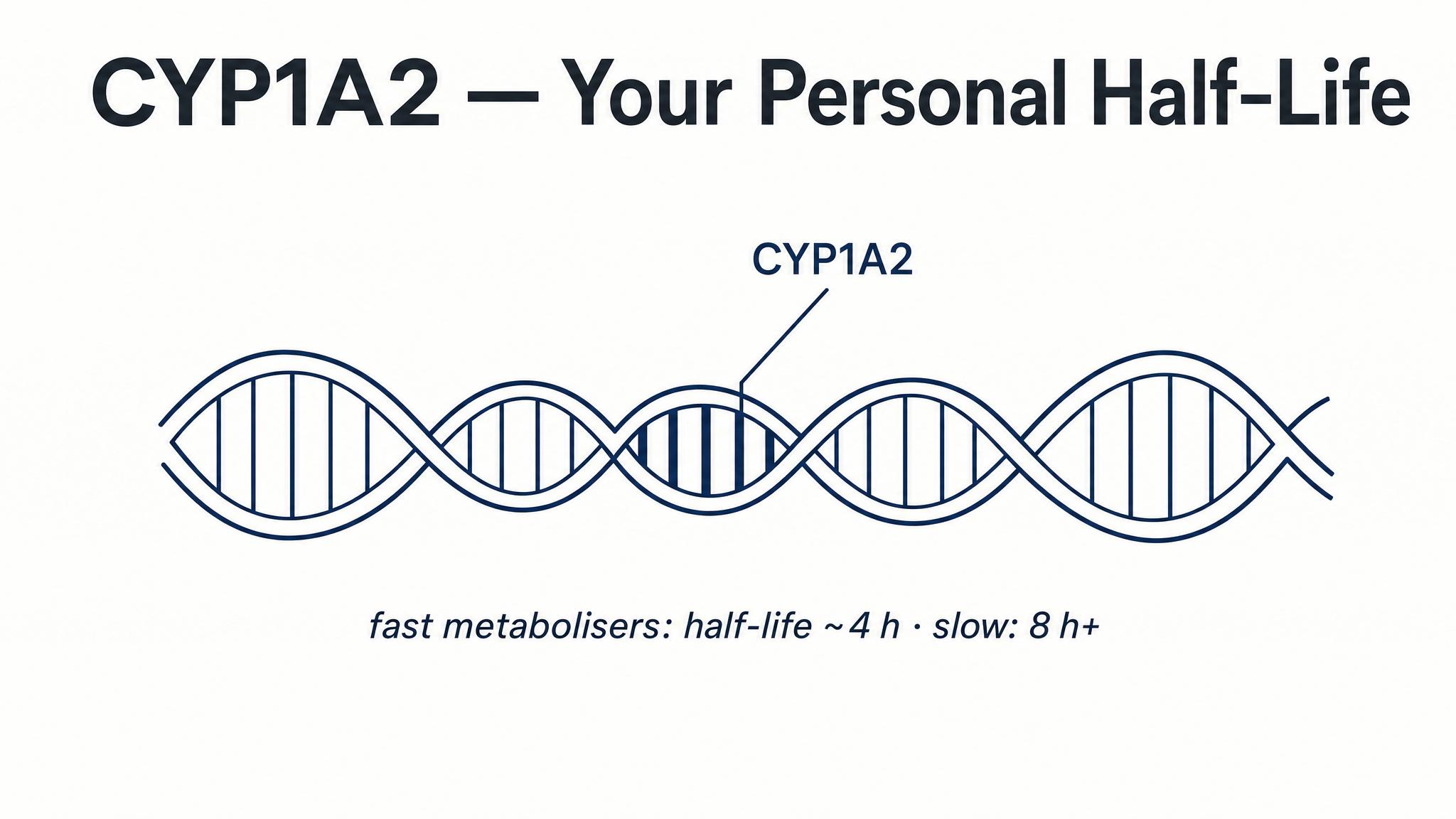




Moving to Sleep — Exercise, Body Temperature, and the Gate to Slumber
This chapter introduces a circadian signal students can feel in their own bodies every day: core body temperature. Students will learn that the SCN drives a daily temperature oscillation — peaking in the late afternoon…
Your gym bro swears late-night workouts help them sleep like a baby. Are they delusional or onto something?
Gym bro science
Do you sleep better in an ice-cold room or cozy-warm? And why do people have such strong opinions about this?
Temperature wars
Have you ever been too pumped up from exercise to fall asleep? Or does working out always knock you out?
Real talk
Would you rather: Wake up at 5am to work out, or squeeze in a workout at 10pm and risk being wired?
Would you rather
Hot take: The whole 'no exercise within 3 hours of bedtime' rule is outdated BS. Agree or disagree?
Hot take
Is taking a cold shower before bed a legit sleep hack or just influencer nonsense?
Hack or hype
Temperature Gate Detective
12-15 minutesStudents track their own peripheral temperature changes in real-time. (1) Have students place hands on neck/forehead to assess warmth, then compare to hands/feet. (2) Discuss with 2-3 neighbors: When do you notice hands/feet warmest vs. coldest during the day? (3) Instructor presents a graph of typical core vs. peripheral temperature rhythms. (4) Students map their personal observations to the vasodilation 'gate to sleep' concept. (5) Volunteers share surprising realizations (e.g., 'My feet are ice cold in late afternoon when I'm most alert!'). (6) Quick instructor synthesis connecting personal experience to the mechanism.
Exercise Timing Myth Trial
15-18 minutes(1) Instructor presents 4 common claims: 'Never exercise within 3 hours of bed', 'Morning exercise is best for sleep', 'Evening yoga helps you sleep', 'Intense evening exercise ruins sleep'. (2) Students stand if they believe a claim, remain seated if skeptical (show of hands alternative). (3) For each claim, select standing and seated students to defend their position (30-60 sec each). (4) Instructor reveals evidence from meta-analyses. (5) Students discuss with neighbors: What surprised you? Why do myths persist? (6) Conclude with evidence-based nuance: intensity, timing, individual differences matter.
Sleep Architecture Scramble
10-12 minutes(1) Instructor projects 8-10 jumbled statements about exercise effects (e.g., 'increases SWS in first half of night', 'reduces REM latency', 'minimal effect on total sleep time', 'decreases sleep onset latency'). (2) Students have 3 minutes to rank these from 'strongest evidence' to 'weakest evidence' individually. (3) Compare rankings with neighbors (2-3 people), discuss discrepancies. (4) Instructor reveals consensus from sleep research, highlighting surprising findings. (5) Quick poll: Which finding most changes how you'll approach exercise?
Temperature Timeline Challenge
8-10 minutes(1) Students receive (or sketch) a blank 24-hour timeline. (2) In pairs with neighbors, plot: core body temperature peak, core temp nadir, typical bedtime, typical wake time, and optimal 'sleep gate' window. (3) Add: When would evening exercise raise core temp? When would effect dissipate? (4) Mark '3-hour rule' window vs. actual evidence-based window. (5) Volunteers share their timelines (project phone photo or sketch on board). (6) Instructor overlays research-based timeline, discussing individual variation and practical implications.
Personal Protocol Designer
15-20 minutes(1) Students receive a realistic case scenario (e.g., 'Alex: shift worker, gym access 6-8pm only, struggles with sleep onset' or 'Jordan: anxious sleeper, loves morning runs, wakes at 5am'). (2) With neighbors, design an evidence-based exercise protocol: timing, intensity, duration, post-exercise cooling strategies. (3) Groups justify choices based on course concepts (temperature gate, sleep architecture, individual factors). (4) Instructor cold-calls 4-5 groups to present 60-second protocols. (5) Class votes on most evidence-based and most creative. (6) Instructor highlights excellent reasoning and common pitfalls.
Vasodilation Visualization
5-8 minutes(1) Instructor asks: 'Why don't we sleep immediately after a hot bath if warmth causes sleep?' (2) Students turn to neighbors, discuss for 90 seconds. (3) Instructor demonstrates with a volunteer: feel forehead vs. hands immediately after imagining hot bath. (4) Key reveal: It's peripheral vasodilation (cooling core via extremities), not overall warmth. (5) Quick challenge: Students explain to neighbors why cold feet keep you awake (vasoconstriction → core temp stays high). (6) Rapid-fire Q&A: students shout out situations that open or close the temperature gate.
Transcript
Picture a winter evening. You've just stepped out of a hot bath, skin flushed and radiating warmth. Within twenty minutes, you feel an almost irresistible heaviness in your eyelids. Or consider the opposite scenario: it's a sweltering August night, your apartment has no air conditioning, and you're lying in bed wide awake at one a-m, sheets kicked off, too hot to sleep. You know intuitively that temperature and sleep are connected, but the relationship is the opposite of what most people assume. It's not warmth that puts you to sleep. It's LOSING warmth.
Now add a second puzzle. Your friend tells you she went for a run at eight p-m and slept like a rock. A fitness blog insists that exercising within four hours of bedtime will ruin your sleep. Who's right? As you'll discover in this chapter, the answer lies in understanding a single elegant mechanism: the rise and fall of your core body temperature, and how both your circadian clock and your behavior, including exercise, baths, and even what you wear on your feet, can shape that curve to open or close what researchers call the temperature gate to sleep.
[short pause]
The Hidden Thermostat: Your Core Body Temperature Rhythm
In Chapter One, we introduced the suprachiasmatic nucleus, or S-C-N, as the master circadian clock. We focused then on its role in timing alertness and melatonin release. But the S-C-N orchestrates another rhythm that you can literally feel if you pay attention: a daily oscillation in CORE BODY TEMPERATURE, or C-B-T, that spans roughly one degree Celsius, about one point eight degrees Fahrenheit, across the twenty-four-hour day.
This oscillation follows a remarkably consistent pattern. Core body temperature begins rising in the early morning hours, typically reaching its peak, the acrophase, in the late afternoon, usually between five and seven p-m for most adults. After this peak, temperature begins a gradual decline that continues through the evening and into the night, reaching its lowest point, the nadir, in the early morning hours, roughly between three and five a-m. As Harding and colleagues found in 2019, this isn't a response to sleeping; it's a proactive signal. Under controlled laboratory conditions where participants stay awake continuously, called a constant routine protocol, the temperature curve persists with the same shape, confirming it's driven by the circadian clock rather than by behavior.
What makes this rhythm so important for sleep is not just correlation, it's causation. The declining phase of the temperature curve is one of the most potent biological triggers for sleep onset. As Kräuchi demonstrated in 2007, sleepiness doesn't simply happen to coincide with cooling; the process of heat loss itself appears to be mechanistically involved in initiating sleep.
[short pause]
Warm Hands, Cool Core: The Vasodilation Mechanism
If the declining temperature curve opens the gate to sleep, what mechanism actually drives that decline? The answer is elegantly simple: your body dumps core heat through your skin, especially through the skin of your hands and feet.
In a landmark study, Kräuchi, Cajochen, Werth, and Wirz-Justice in 2000 measured multiple physiological variables simultaneously under constant-routine conditions and tested which was the best predictor of how quickly a person fell asleep. They examined core body temperature, heart rate, melatonin onset, subjective sleepiness ratings, and something called the DISTAL-PROXIMAL SKIN TEMPERATURE GRADIENT, or D-P-G, the difference in temperature between the extremities, hands and feet, and the trunk. The result was striking: the D-P-G was the single strongest predictor of sleep-onset latency, outperforming every other variable.
Here's what's happening physiologically. In the hours before sleep, the S-C-N signals a process called peripheral vasodilation. The blood vessels in your hands, feet, and face dilate, flooding these extremities with warm blood. You can feel this: your hands and feet get warmer in the evening. This warmth at the surface isn't keeping heat in; it's radiating heat out, like opening the vents on an overheated engine. As heat pours off your extremities, your core temperature drops. The greater this distal vasodilation, the warmer your hands and feet relative to your trunk, the faster you fall asleep, as Kräuchi and colleagues found in 2000.
This process begins approximately one to two hours before habitual sleep onset, as Harding and colleagues reported in 2019, which means your body is actively preparing for sleep well before you feel subjectively drowsy. It also means that anything interfering with this heat dissipation, cold feet constricting blood vessels, a room that's too warm preventing the core-to-surface gradient, can delay sleep onset even if you're otherwise tired.
[short pause]
Recall from Chapter Four how adenosine builds up across the day to create homeostatic sleep pressure, Process S. Now consider: if Process S represents chemical sleep pressure and the temperature decline represents a circadian sleep gate, what happens when these two signals are misaligned, say, high adenosine pressure but a still-rising temperature curve? How might that feel experientially?
This vasodilation mechanism also explains a folk remedy that turns out to be scientifically sound: wearing socks to bed. A Swiss study found that warming the feet accelerated vasodilation and shortened sleep-onset latency. It's not the warmth itself that matters, it's that warm feet mean dilated blood vessels, which means faster core heat loss, which means the temperature gate opens sooner.
[short pause]
The Warm Bath Paradox: Heat In, Sleep Out
With the temperature-gate model established, we can now solve a genuine puzzle: why does a warm bath or shower before bed help people fall asleep faster? At first glance, it seems contradictory. If cooling promotes sleep, why would adding heat help?
The answer is that a warm bath doesn't just add heat, it strategically redistributes it. When you immerse yourself in warm water, forty to forty-two point five degrees Celsius, or one hundred four to one hundred eight point five degrees Fahrenheit, blood rushes to the surface of your skin throughout your body, dramatically increasing peripheral vasodilation. When you step out of the bath, all of those dilated blood vessels are now exposed to cooler ambient air, creating a rapid and exaggerated core temperature drop, steeper than what would have occurred naturally, as Haghayegh and colleagues found in 2019.
Haghayegh and colleagues in 2019 conducted a systematic review and meta-analysis of seventeen studies and found that passive body heating scheduled one to two hours before bedtime significantly shortened sleep-onset latency by an average of about ten minutes and improved both sleep efficiency and subjective sleep quality. The optimal timing, one to two hours before bed, aligns perfectly with the temperature-gate model: the bath amplifies the vasodilation that the S-C-N is already initiating, supercharging the natural cooling process.
[short pause]
As adapted from Kräuchi in 2007: It is the rate of heat loss, not warmth itself, that the sleeping brain responds to. The warm bath is a delivery mechanism for rapid cooling, a physiological trick that hijacks the circadian thermoregulatory system.
[short pause]
Exercise and Sleep: What the Evidence Actually Shows
Now we arrive at one of the most powerful, and most misunderstood, tools for improving sleep: physical exercise. The popular health message is simple: exercise helps you sleep. And broadly speaking, this is true. But the details matter enormously, and the oft-repeated warning to never exercise in the evening turns out to be far more nuanced than most people realize.
The Big Picture: Exercise Improves Sleep Architecture
Kredlow, Capozzoli, Hearon, Calkins, and Otto in 2015 conducted a comprehensive meta-analysis of sixty-six studies examining the relationship between physical activity and sleep. Their findings were clear: regular exercise produces moderate beneficial effects on overall sleep quality. Specifically, regular exercisers showed improvements in sleep-onset latency, total sleep time, sleep efficiency, and, notably, N-three SLOW-WAVE SLEEP, the deep restorative stage we discussed in Chapter Two.
Yamanaka and colleagues in 2021 added important mechanistic detail, showing that vigorous exercise, sixty minutes at sixty percent V-O-two-max, significantly increased delta power during N-three sleep and improved slow-wave stability, as measured by E-E-G, or electroencephalography. In other words, exercise doesn't just give you more deep sleep, it gives you better quality deep sleep, with larger, more stable slow waves.
[short pause]
Why Exercise Improves Sleep: Three Converging Mechanisms
Exercise improves sleep through at least three mechanisms, each connecting back to concepts we've already explored:
First, the TEMPERATURE PATHWAY. Exercise raises core body temperature by one to two degrees Celsius. In the hours following exercise, C-B-T drops, often overshooting slightly below baseline. This post-exercise cooling mimics and amplifies the circadian temperature decline, potentially opening the temperature gate to sleep in the same way a warm bath does, as Harding and colleagues found in 2019.
Second, the ADENOSINE PATHWAY, or Process S. Remember from Chapter Four that adenosine accumulates in the brain during wakefulness, building homeostatic sleep pressure. Dworak, McCarley, Kim, Kalinchuk, and Basheer in 2007 demonstrated in animal studies that intense exercise increases brain adenosine concentrations to two hundred twenty-nine percent of resting levels, essentially turbocharging Process S. This may be one reason why people who exercise regularly report feeling more genuinely tired at bedtime rather than the wired-but-exhausted state of sedentary individuals.
Third, the CIRCADIAN PATHWAY. When exercise occurs outdoors, it delivers bright-light exposure, the single most powerful zeitgeber we discussed in Chapter Three. A morning jog, therefore, is doing double duty: elevating core temperature, which will later decline, and delivering the light signal that anchors your circadian clock. This is why morning outdoor exercise may be the single most potent behavioral intervention for sleep health.
[short pause]
Given what you learned in Chapter Three about light's effects on the circadian clock, and what you now know about the temperature pathway, can you predict why exercising outdoors in the morning might be more beneficial for sleep than exercising indoors at the same time? How many mechanisms overlap?
[short pause]
The Evening Exercise Myth: What the Evidence Really Says
Now for the part that surprises most students. The widespread advice to avoid exercise in the evening, typically framed as no exercise within three to four hours of bedtime, is substantially more nuanced than popular health media suggests. The critical study here is the systematic review and meta-analysis by Stutz, Eiholzer, and Spengler in 2019, which examined twenty-three studies on evening exercise and sleep.
Their findings challenge the blanket prohibition:
First, evening exercise does NOT harm sleep for most people. Across studies, evening exercise actually increased slow-wave sleep and decreased light stage one sleep compared to no-exercise controls, meaning it improved sleep depth.
Second, the exception is narrow and specific. Sleep-onset latency, total sleep time, and sleep efficiency were impaired only after vigorous-intensity exercise ending less than one hour before bedtime. Moderate-intensity evening exercise, and vigorous exercise ending two or more hours before bed, showed no negative effects.
Third, for many people, evening exercise is beneficial. The post-exercise temperature drop, occurring sixty to ninety minutes after a workout, can actually facilitate the natural temperature decline and open the sleep gate.
[short pause]
The practical implication is important: if the only time you can exercise is in the evening, do it. The sleep benefits of regular exercise almost certainly outweigh any small risk from timing. The one scenario to approach cautiously is an all-out sprint session or high-intensity interval workout finishing right at bedtime, and even then, individual variation is significant. Some people sleep beautifully after intense evening exercise; others don't. Your own body's response matters more than a generic rule.
[short pause]
Putting It All Together: Exercise Timing and Your Circadian Window
Rather than memorizing rigid rules about when to exercise, the more powerful approach is to understand the underlying mechanisms and apply them to your own life. Here's a framework for thinking about exercise timing relative to the circadian signals we've discussed:
Morning Exercise, six to ten a-m. Morning exercise, especially outdoors, is a circadian powerhouse. It delivers bright-light exposure that strengthens S-C-N entrainment, as we learned in Chapter Three, elevates core temperature early in the day, reinforcing the natural temperature rise, and generates adenosine that will compound with the day's normal accumulation, as discussed in Chapter Four. For people who struggle with sleep-onset timing or who have delayed circadian tendencies, morning exercise is arguably the single best behavioral intervention available.
Afternoon Exercise, two to five p-m. Exercise in the mid-to-late afternoon coincides with the peak of the temperature curve. Vigorous exercise at this time produces the largest absolute temperature elevation, and the subsequent post-exercise cooling aligns naturally with the evening temperature decline. Many studies show that afternoon exercise produces the greatest improvements in N-three deep sleep, as Kredlow and colleagues found in 2015.
Evening Exercise, six to nine p-m. Moderate evening exercise is safe and beneficial for the vast majority of people. The post-exercise temperature drop, occurring roughly sixty to ninety minutes later, may actually facilitate sleep onset by amplifying the natural circadian cooling. The key exception: vigorous exercise ending less than one hour before your intended bedtime may delay sleep onset for some individuals, likely due to elevated sympathetic nervous system activation and a still-elevated core temperature that hasn't had time to decline, as Stutz and colleagues reported in 2019.
[short pause]
Imagine two students: one is a strong morning chronotype who wakes at six a-m and sleeps at ten p-m; the other is a late chronotype who wakes at ten a-m and sleeps at two a-m. Evening exercise at eight p-m falls at very different points in their respective circadian curves. How might the same workout affect them differently?
[short pause]
The Integrated View: Building Your Sleep-Promoting Day
By now you've accumulated an impressive toolkit of behavioral levers that affect sleep. From Chapter Three, you understand how light exposure entrains your circadian clock. From Chapter Four, you know how caffeine blocks adenosine and how timing your last cup of coffee matters. And from this chapter, you've learned how exercise and temperature regulation interact with both Process C and Process S.
The most powerful insight is that these interventions don't operate in isolation, they COMPOUND. A morning outdoor jog delivers light exposure and temperature elevation and adenosine generation simultaneously. Conversely, negative behaviors can compound too: caffeine at five p-m blocks the adenosine signal, bright screen light at ten p-m suppresses melatonin and delays the circadian temperature decline, and a vigorous gym session at ten-thirty p-m elevates core temperature right when it should be falling. Each alone might be manageable; together, they can create a perfect storm of insomnia.
The goal isn't to follow a rigid protocol, it's to understand the principles well enough to make informed choices that fit your life. A student who can only exercise at eight p-m isn't doomed to bad sleep. But that student might choose moderate intensity over high intensity, might cut caffeine by early afternoon, and might take a warm shower ninety minutes before bed to maximize the temperature drop. Understanding the mechanisms turns rigid rules into flexible strategies.
[short pause]
To summarize the KEY concepts from this chapter:
Core body temperature follows a circadian rhythm, peaking in the late afternoon and reaching its nadir in the early morning, and the declining phase of this curve is a potent biological trigger for sleep onset.
Sleep onset is best predicted by the distal-proximal skin temperature gradient: warm hands and feet, vasodilation, radiate heat outward, cooling the core and opening the temperature gate to sleep.
A warm bath one to two hours before bed works not because of the warmth, but because it amplifies peripheral vasodilation and accelerates the subsequent core temperature drop.
Regular exercise improves sleep quality, increases N-three deep sleep, and reduces sleep-onset latency through three converging mechanisms: post-exercise cooling, increased adenosine accumulation, and when outdoors, circadian light exposure.
The blanket advice to avoid evening exercise is overstated. Evening exercise is neutral or beneficial for most people; only vigorous exercise ending less than one hour before bedtime shows potential to delay sleep onset.
Morning outdoor exercise may be the most powerful single behavioral intervention for sleep health, delivering simultaneous benefits through the temperature, adenosine, and circadian pathways.
Sleep interventions compound, positively and negatively. Understanding the mechanisms lets you combine light, caffeine timing, and exercise strategically rather than following rigid rules.
[short pause]
Looking Ahead
You now understand three major levers for sleep: light, caffeine, and exercise-temperature. In Chapter Six, we'll turn to the dimension of sleep that most directly affects your nightly experience, the sleep environment. Temperature, noise, and darkness aren't just preferences; they're physiological requirements. We'll explore the surprising science of bedroom temperature, hint: it connects directly to the vasodilation mechanisms from today, why certain sounds disrupt sleep architecture even when you don't wake up, and how to engineer your sleeping space for optimal recovery. Your bedroom is either working for your sleep or against it, and most students' bedrooms are working against them.

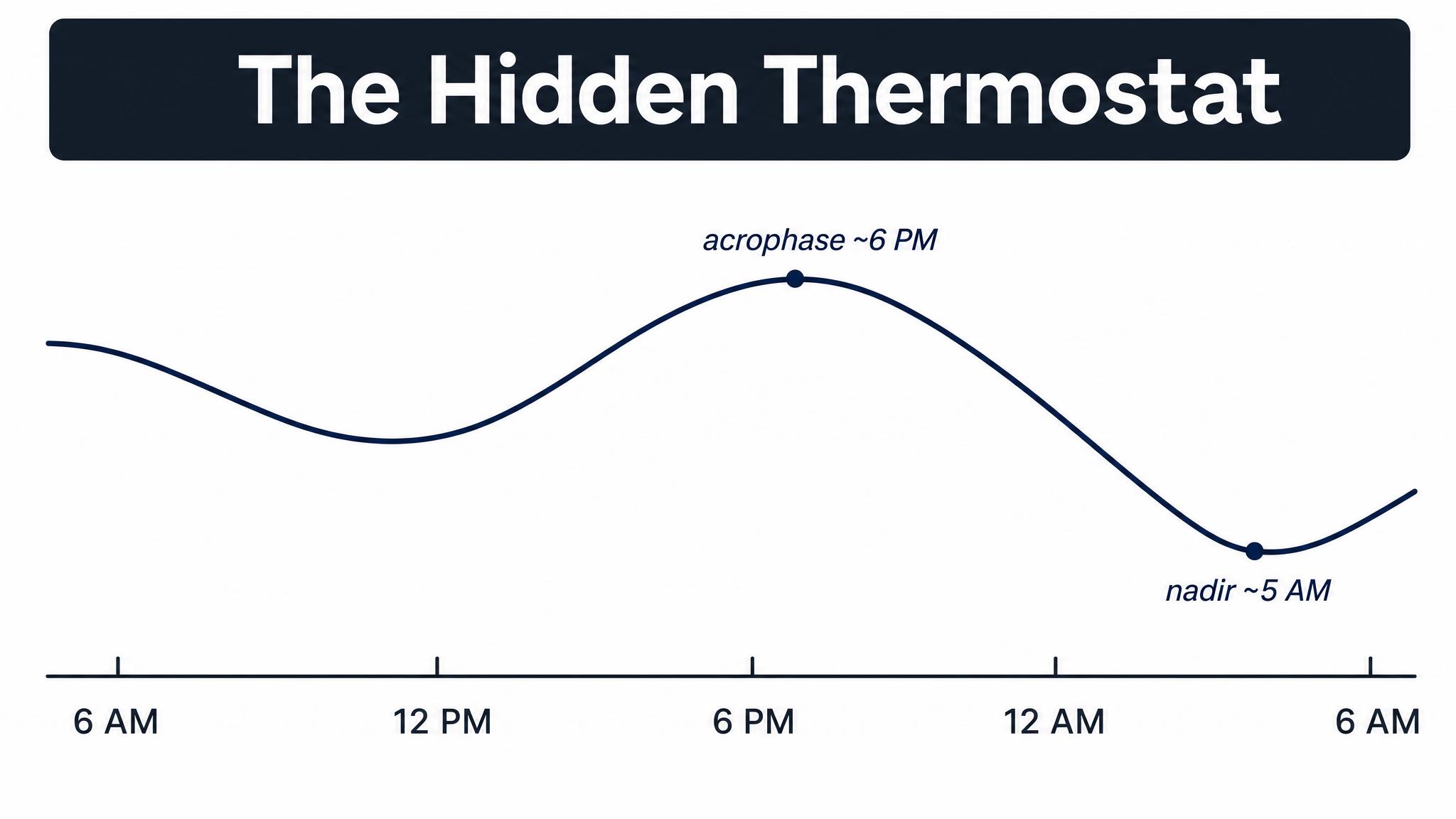

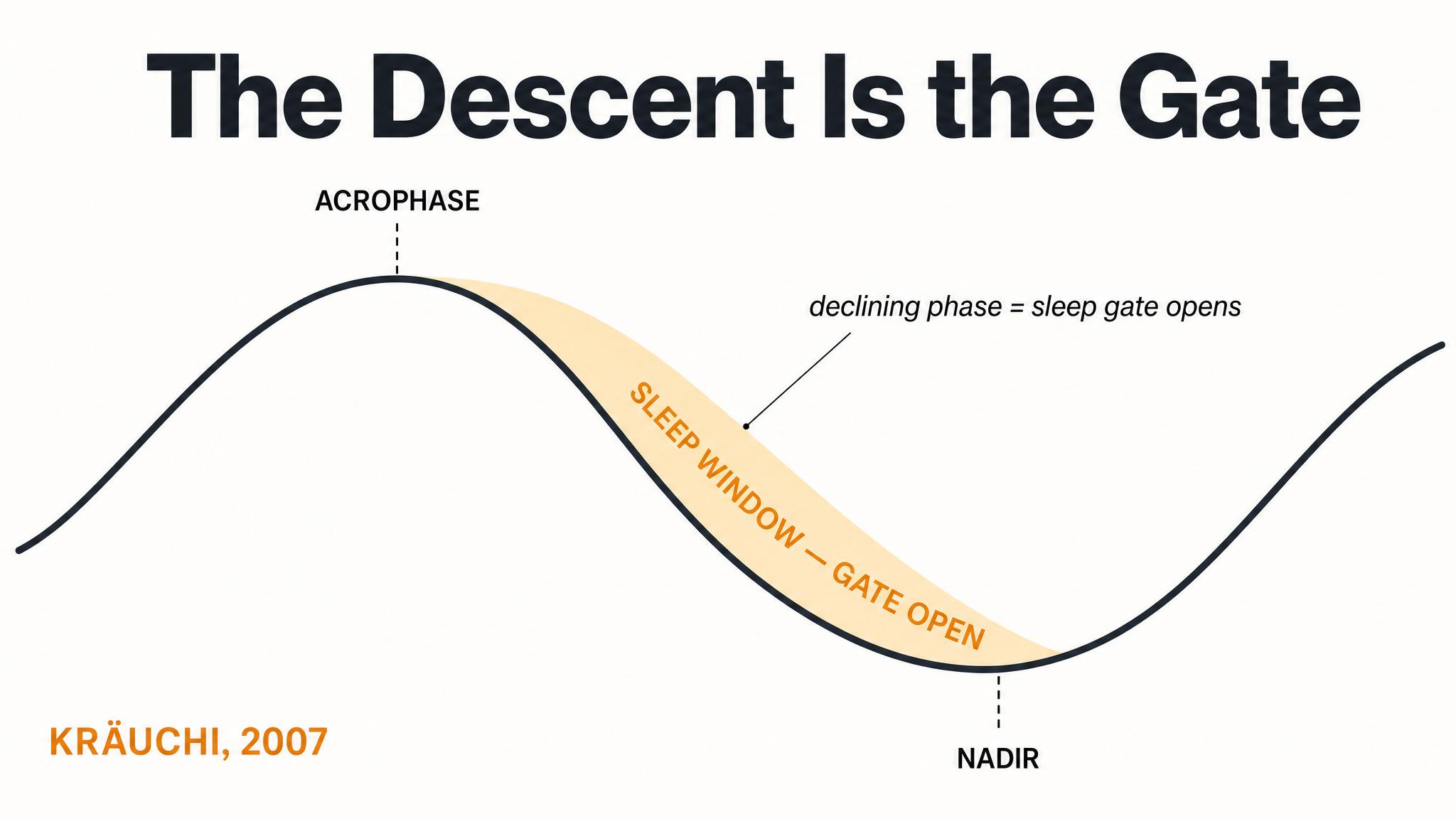


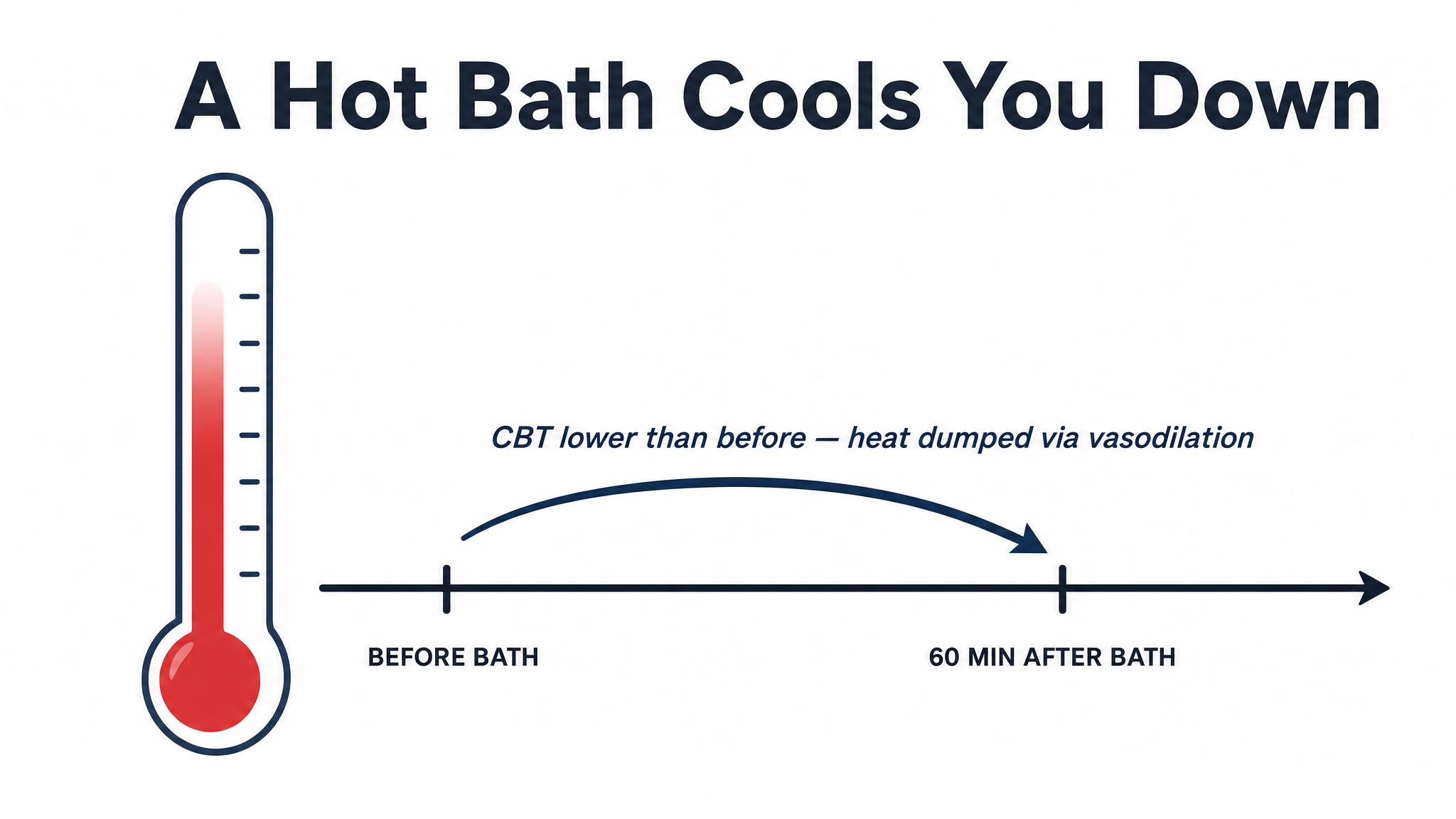

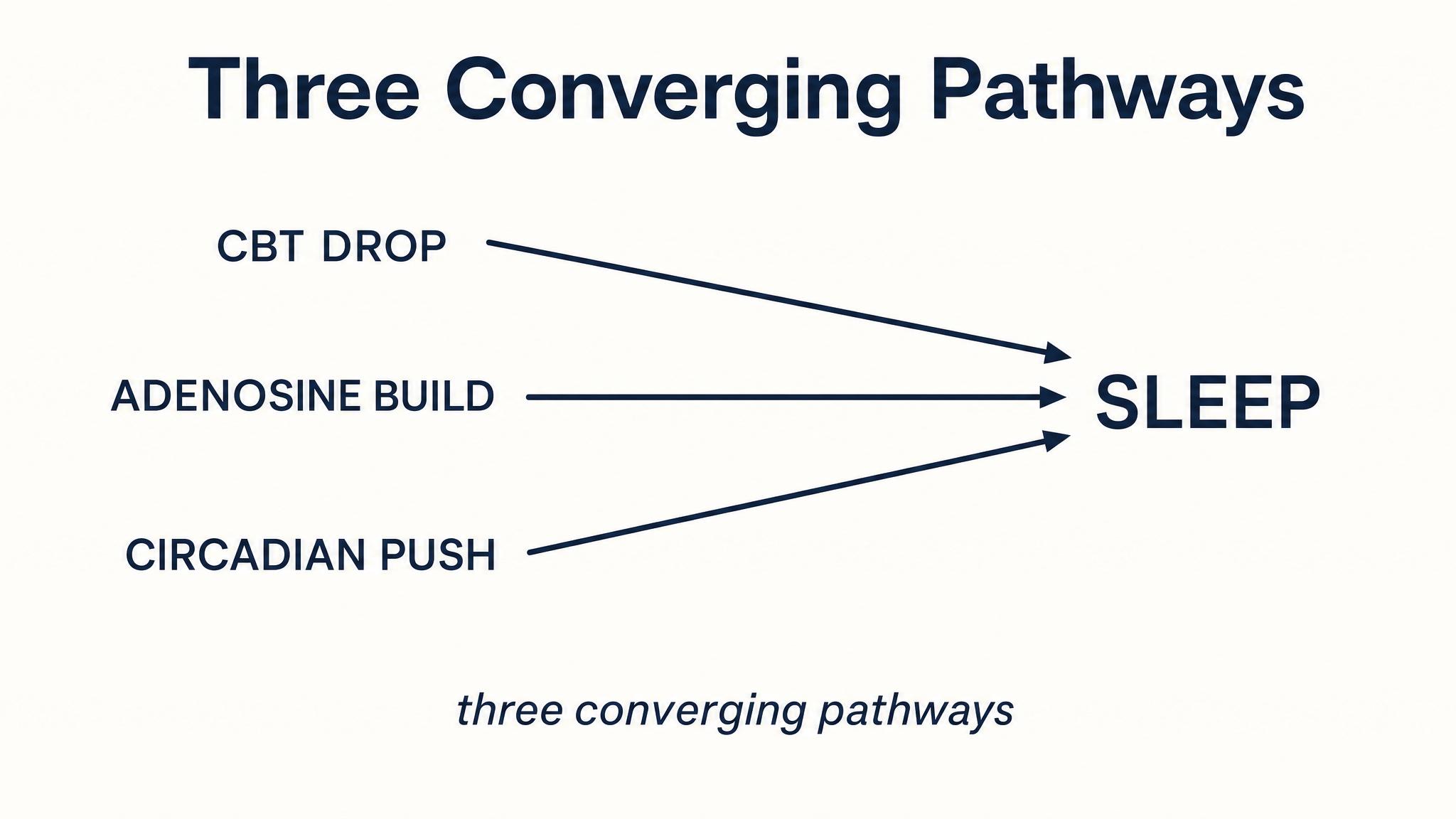



Screens, Stimulation, and Wind-Down — Why Your Brain Won't Shut Up
This is the chapter where the course challenges one of the most widely repeated pieces of sleep advice: 'blue light from screens is ruining your sleep.' With three chapters of photobiology, pharmacology, and physiology…
What's more likely to wreck your sleep: doomscrolling TikTok or binge-watching a chill show on Netflix? Why?
Which is worse
Be honest: have you EVER actually stopped looking at screens an hour before bed like you're supposed to? What happened?
Real talk
Would you rather give up all screens after 9pm for a month, or give up coffee/energy drinks for a month?
Would you rather
Hot take: It's not the blue light keeping you awake — it's that you're watching something way more interesting than your own thoughts. Agree or nah?
Hot take
What's your worst pre-bed screen habit? Checking work emails? Instagram stalking? Online shopping? Reading Twitter arguments?
Confess yours
Do you think your brain actually cares whether you're scrolling on your phone vs. reading a physical book before bed, or is that just something wellness influencers made up?
Myth or truth
Transcript
It's eleven twenty-three PM. You told yourself you'd be asleep by eleven. Your phone is in your hand — you're not sure exactly when you picked it up, but you're three articles deep into a news rabbit hole, your heart rate is slightly elevated from a comment section argument you almost joined, and the blue-light-filtering app you installed last month is dutifully tinting your screen orange. You're doing the "right" thing about the light. So why do you feel more awake now than you did an hour ago?
Here's the uncomfortable truth that most sleep advice glosses over: the orange tint on your screen is addressing the smaller problem. The real reason your brain won't shut up has almost nothing to do with wavelengths of light — and almost everything to do with what's happening between your ears. This chapter will recalibrate what you think you know about screens and sleep, and replace vague guilt with a precise, evidence-based strategy.
[short pause]
If you've encountered any sleep advice in the last decade, you've almost certainly heard some version of this claim: the blue light from your phone suppresses melatonin and ruins your sleep. It's become one of the most widely repeated pieces of health advice in modern culture. It sells blue-light-blocking glasses, night-mode software, and amber screen protectors. And it's not entirely wrong — but it's dramatically overstated.
Let's revisit what we learned earlier. MELANOPSIN, the photopigment in your intrinsically photosensitive retinal ganglion cells, is indeed most sensitive to light in the short-wavelength "blue" range, peaking around four hundred and eighty nanometers. When melanopsin detects sufficient light, it sends signals to the suprachiasmatic nucleus that suppress melatonin production. Screens do emit light in this melanopsin-sensitive range. All of that is accurate.
The critical question, however, is one of dose. How much light are we actually talking about?
[short pause]
The landmark study by Gooley and colleagues in two thousand and eleven demonstrated that exposure to room light of approximately two hundred LUX before bedtime suppressed melatonin onset in ninety-nine percent of participants and shortened melatonin duration by roughly ninety minutes. This study is frequently cited as evidence against screen use — but notice the exposure level. Two hundred lux is the illumination of a moderately well-lit living room, not a phone screen at arm's length.
What does your phone actually produce? Technical analyses have consistently found that a smartphone at typical viewing distance, approximately thirty to forty centimeters, delivers between twenty and eighty lux at eye level, depending on brightness settings and ambient conditions, as Oh and colleagues reported in two thousand and fifteen, and as Huiberts and colleagues confirmed in two thousand and twenty-two. Oh and colleagues measured melatonin suppression values of only seven to eleven percent for typical smartphone use in a dark room — a far cry from the near-total suppression found at two hundred plus lux. Spitschan and colleagues in two thousand and twenty-three confirmed this dose-dependent relationship in a controlled study of seventy-two participants, showing that while smartphone-level light at around twenty lux did produce measurable effects on sleep latency, the magnitude increased substantially with higher melanopic irradiance levels characteristic of tablets at around forty lux and computer screens at around eighty lux.
The systematic review by Shechter and colleagues in two thousand and twenty reinforced this picture: the studies showing robust melatonin suppression predominantly used light exposures at or above two hundred lux. Below that threshold, effects were present but considerably more modest. Huiberts and colleagues noted in their field study that screen illuminance at eye level in typical seated positions usually remains below fifty lux — well below the levels where dramatic melatonin suppression has been consistently documented.
[short pause]
Before we continue, consider this: if the blue light from a phone screen at arm's length produces roughly forty to eighty lux, and robust melatonin suppression in studies typically requires two hundred plus lux, what else might explain why scrolling your phone at midnight makes it so hard to fall asleep?
To be clear, this is not permission to blast yourself with screen light until midnight. The effects at lower lux levels are real — they're just more modest than the cultural narrative suggests. A phone screen in a dark room at full brightness is not nothing. But framing blue light as the primary mechanism by which screens disrupt sleep is, as the evidence stands, a significant overstatement. There is a larger, more potent mechanism at work.
[short pause]
If you've ever tried to fall asleep after a heated text exchange, an anxiety-inducing news scroll, or a tense final round in an online game, you already have intuitive evidence for what the research now strongly supports: the content on your screen matters far more than the light coming off of it.
Kalmbach and colleagues in two thousand and twenty conducted polysomnographic recordings of fifty-two adults across multiple nights and found that nocturnal COGNITIVE AROUSAL — the racing, ruminative mental activity that keeps your mind churning — was consistently and robustly associated with objective sleep disturbance. Crucially, cognitive arousal was more strongly linked to difficulty falling asleep than clinical insomnia diagnosis, depression, or even somatic, that is body-based, arousal. Your mind racing is, in measurable physiological terms, more disruptive to sleep onset than your body being tense.
Exelmans and Van den Bulck in two thousand and seventeen explored this in the specific context of screen use, studying four hundred and twenty-three young adults and their binge-viewing habits. Their finding was telling: binge-watching before bed negatively affected sleep quality, and the mechanism was not light exposure but increased cognitive pre-sleep arousal. The engagement with narrative content — the suspense, the emotional investment, the "just one more episode" pull — was what kept the brain activated.
Hale and Guan in two thousand and fifteen synthesized the broader literature and identified three distinct pathways through which screens disrupt sleep: time displacement, where screens simply replace sleep time, light exposure effects, and psychological and physiological arousal from content. Their review concluded that arousal may be the most important pathway, mediating sleep problems even with non-violent, seemingly innocuous media use.
The National Sleep Foundation's consensus statement, as reported by Bartel and colleagues in two thousand and twenty-four, affirmed this multi-pathway model, noting that screen content evokes psychological consequences — fear, anxiety, excitement, social comparison — that drive cognitive arousal, which then interacts with light emission to delay and disrupt sleep. The panel specifically noted that the arousal pathway applies across all ages and is not limited to obviously stimulating content.
[short pause]
When we say screens cause "arousal," we're not using the word loosely. The SYMPATHETIC NERVOUS SYSTEM — your fight-or-flight infrastructure — activates in response to stimulating content. Heart rate increases slightly. Cortisol, which should be declining in the evening, gets a nudge. The prefrontal cortex remains engaged in evaluative processing. The default mode network, which normally begins to quiet during the transition toward sleep, stays active as you mentally compose responses, evaluate social comparisons, or anticipate what happens next in a show.
This brings us to a fundamental requirement of falling asleep that most people never think about explicitly: your nervous system must make a transition.
[short pause]
Falling asleep is not like flipping a switch. It's more like slowly easing a car from highway speed down to a stop — and it requires specific physiological shifts that take time and the right conditions.
Bonnet and Arand in two thousand and ten demonstrated that the transition from wakefulness to sleep is characterized by a progressive shift from sympathetic to PARASYMPATHETIC nervous system dominance. In normal sleepers, this shift happens gradually: heart rate slows, blood pressure drops, muscle tension decreases, core body temperature falls, and cortisol levels decline. Participants with insomnia in their study showed a revealing pattern — constant sympathetic hyperactivation that persisted even after sleep onset, resulting in lighter, less restorative sleep.
Research on autonomic activity during sleep, as described by Okon-Singer and colleagues in two thousand and twenty, has confirmed that successful sleep transition requires the withdrawal of sympathetic activity and increasing predominance of parasympathetic activity. When you're scrolling through content that triggers emotional reactions — outrage at a news article, anxiety about a work email, competitive intensity from a game — you are actively opposing this necessary transition. You're pressing the accelerator while trying to brake.
[short pause]
Recall how the suprachiasmatic nucleus coordinates the timing of melatonin release with declining light signals. Now consider: even if melatonin is being released on schedule, can you fall asleep easily if your sympathetic nervous system is highly activated? What does this tell you about the relative importance of hormonal timing versus nervous system state?
[short pause]
This understanding leads to a crucial distinction that blanket "no screens before bed" advice misses entirely. Consider these two scenarios:
Scenario A: You're reading a familiar novel on a Kindle set to low brightness with warm-tone backlighting, in a dimly lit room. Your breathing is slow. You're feeling pleasantly drowsy.
Scenario B: You're scrolling Twitter on your phone at medium brightness, encountering a series of posts about a political controversy. Your jaw is slightly clenched. You've composed and deleted two replies.
Both involve "screen time." Both emit some light in the melanopsin-sensitive range. But the physiological states they produce are vastly different. Scenario A involves minimal cognitive arousal and modest light exposure. Scenario B involves significant sympathetic activation, cortisol elevation, and sustained prefrontal engagement — all of which actively oppose the sleep transition, regardless of what night mode your phone is in.
This is the reframing this chapter offers: the question is not simply "are you using a screen?" but "is this activity moving your nervous system toward sleep or away from it?"
[short pause]
If the brain needs time to shift from sympathetic to parasympathetic dominance, the practical implication is clear: you need a WIND-DOWN BUFFER ZONE — a period of progressively declining stimulation in the sixty to ninety minutes before your intended sleep time.
Irish and colleagues in two thousand and fifteen reviewed behavioral strategies for addressing insomnia and recommended establishing a bedtime routine thirty to forty-five minutes before sleep to help transition the brain by "winding down and slowing central nervous system activity." Their review emphasized that consistency matters: when the same sequence of low-stimulation activities precedes sleep night after night, the brain begins to treat that sequence as a conditioned cue — a signal that the sleep transition should begin. Think of it like Pavlov's dogs, but instead of a bell triggering salivation, a consistent wind-down routine triggers the neurochemical cascade that precedes sleep.
The structure of an effective wind-down buffer zone follows a simple principle: PROGRESSIVE DE-ESCALATION. Arousal should trend downward across the period. Light exposure should decline. Cognitive demands should decrease. Physical activity should shift from active to restful. This doesn't require rigid rules or complete abstinence from all stimulation — it requires a direction of travel.
[short pause]
A practical wind-down protocol addresses both the moderate light concern and the more significant arousal concern. Consider these evidence-based elements:
Light management, which is moderate priority: Dim overhead lights. If using screens, reduce brightness and use warm-tone settings — not because this eliminates the light effect, but because it reduces it meaningfully at minimal effort.
Arousal management, which is high priority: Transition from high-engagement activities to low-engagement ones. Replace stimulating content with calming content. Avoid emotional triggers — work email, news, social media arguments.
Routine consistency, also high priority: Perform the same sequence of activities in the same order. This builds conditioned associations between these behaviors and sleep onset.
Body temperature: A warm bath or shower sixty to ninety minutes before bed accelerates the core body temperature drop that accompanies sleep onset.
Cognitive offloading: Journaling or a brief to-do list for tomorrow can reduce the cognitive intrusions that fuel pre-sleep rumination.
[short pause]
If you've ever felt frustrated at yourself for failing to stop scrolling when you know you should be sleeping, here's something important: it's not a character flaw. It's neurochemistry working exactly as predicted.
Recall that adenosine accumulates across the day as part of Process S, the homeostatic sleep drive. By late evening, adenosine levels are high, creating the pressure that makes you feel sleepy. But adenosine doesn't just make you drowsy — it also impairs the brain region most responsible for impulse control, planning, and resisting temptation: the PREFRONTAL CORTEX.
The prefrontal cortex is disproportionately sensitive to fatigue. When adenosine levels are high and Process S pressure is strong, the very brain region you need to make the decision to put the phone down is operating at reduced capacity. You are, in effect, relying on the weakest version of your willpower to resist some of the most compelling stimulation modern technology has ever produced. Social media platforms, news sites, and video streaming services are explicitly engineered by teams of designers to maximize engagement — to make "one more scroll" or "one more episode" feel irresistible. Pitting a fatigued prefrontal cortex against billion-dollar engagement algorithms is not a fair fight.
[short pause]
If your prefrontal cortex is already impaired by high adenosine at eleven PM, what does this suggest about when you should be making your "phone down" decision? Is the moment of temptation the right time to exercise self-control?
[short pause]
The solution, consistent with decades of behavioral science, is PRE-COMMITMENT — making the decision earlier in the day, when your prefrontal cortex is still functioning well, and designing your environment so that the desired behavior becomes the path of least resistance at night.
Effective pre-commitment strategies include:
Environmental design: Charging your phone in another room. If you use your phone as an alarm, buy a simple alarm clock — it may be the best ten pounds you ever spend on sleep.
Automated limits: Setting app timers that enforce breaks after a certain time. Many phones now include "bedtime mode" that greyscales the screen and limits notifications.
Friction engineering: Adding steps between you and the stimulating content. Logging out of social media apps so re-engaging requires deliberate effort. Removing the most tempting apps from your home screen after nine PM.
Replacement, not removal: Having a specific, enjoyable, low-arousal alternative ready — a physical book on the nightstand, a calm podcast queued up — so that you're not left with a void that willpower must fill.
The logic is straightforward: don't fight adenosine. Plan around it.
[short pause]
Let's synthesize what we've covered into a clear, evidence-based framework for thinking about screens and sleep:
Blue light from screens is a real but modest concern. At typical viewing distances, phones produce twenty to eighty lux — below the two hundred plus lux threshold for robust melatonin suppression. Dimming your screen and using warm-tone modes is sensible, easy, and worth doing. But it's not the main event.
Cognitive and emotional arousal is the dominant mechanism. Stimulating content activates the sympathetic nervous system, sustains cortical arousal, and directly opposes the parasympathetic shift necessary for sleep onset. This applies regardless of blue-light filters.
Not all screen use is equivalent. A calm podcast or an e-reader novel at low brightness is fundamentally different from competitive gaming or doomscrolling. The cognitive context matters more than the wavelength.
A wind-down buffer zone of sixty to ninety minutes is your most powerful tool. Use it to progressively decrease arousal, dim your environment, and perform consistent pre-sleep routines that become conditioned cues for the sleep transition.
Pre-commitment strategies outperform willpower. Design your environment earlier in the day when your prefrontal cortex can actually make good decisions.
This reframing is both scientifically honest and practically liberating. Rather than anxiously avoiding all screens — an increasingly unrealistic expectation in modern life — you can make informed decisions about which screen activities to avoid near bedtime and why. You can stop buying expensive blue-light-blocking glasses and invest that energy instead in redesigning your last ninety minutes. And perhaps most importantly, you can stop blaming yourself when willpower fails at eleven PM and start building systems that make the right choice the easy choice.
The goal is not to be anti-screen. The goal is to be pro-transition — to give your nervous system the declining gradient of stimulation it needs to shift from wakefulness into sleep.
[short pause]
Let's review the key points. Blue light from phone and tablet screens at typical viewing distances, that's twenty to eighty lux, falls well below the two hundred plus lux threshold for robust melatonin suppression shown in controlled studies — the concern is real but overstated.
Cognitive and emotional arousal from screen content — not the light itself — is the primary mechanism by which evening screen use disrupts sleep, activating the sympathetic nervous system and opposing the transition to sleep.
Not all screen use is equal: low-arousal screen activities like calm podcasts or e-readers at low brightness are fundamentally different from high-arousal ones like social media scrolling, news feeds, or competitive games in their sleep impact.
A sixty to ninety minute wind-down buffer zone of progressively declining stimulation, dim lighting, and consistent pre-sleep routines creates conditioned cues that help the brain initiate the wake-to-sleep transition.
High adenosine levels at bedtime impair the prefrontal cortex, making willpower-based decisions to stop scrolling unreliable — pre-commitment strategies and environmental design are more effective than relying on tired decision-making.
The sympathetic-to-parasympathetic nervous system shift is a necessary precondition for sleep onset; any activity that sustains sympathetic activation — regardless of its light properties — will oppose this transition.
[short pause]
Now that you understand how to manage the final hours before sleep, the next chapter turns to the sleep environment itself. What does your bedroom need to be — in terms of temperature, sound, light, and psychological associations — to support the fragile transition you've just learned to protect? We'll explore the science of the ideal sleep environment and discover why your bedroom may be unconsciously working against you.



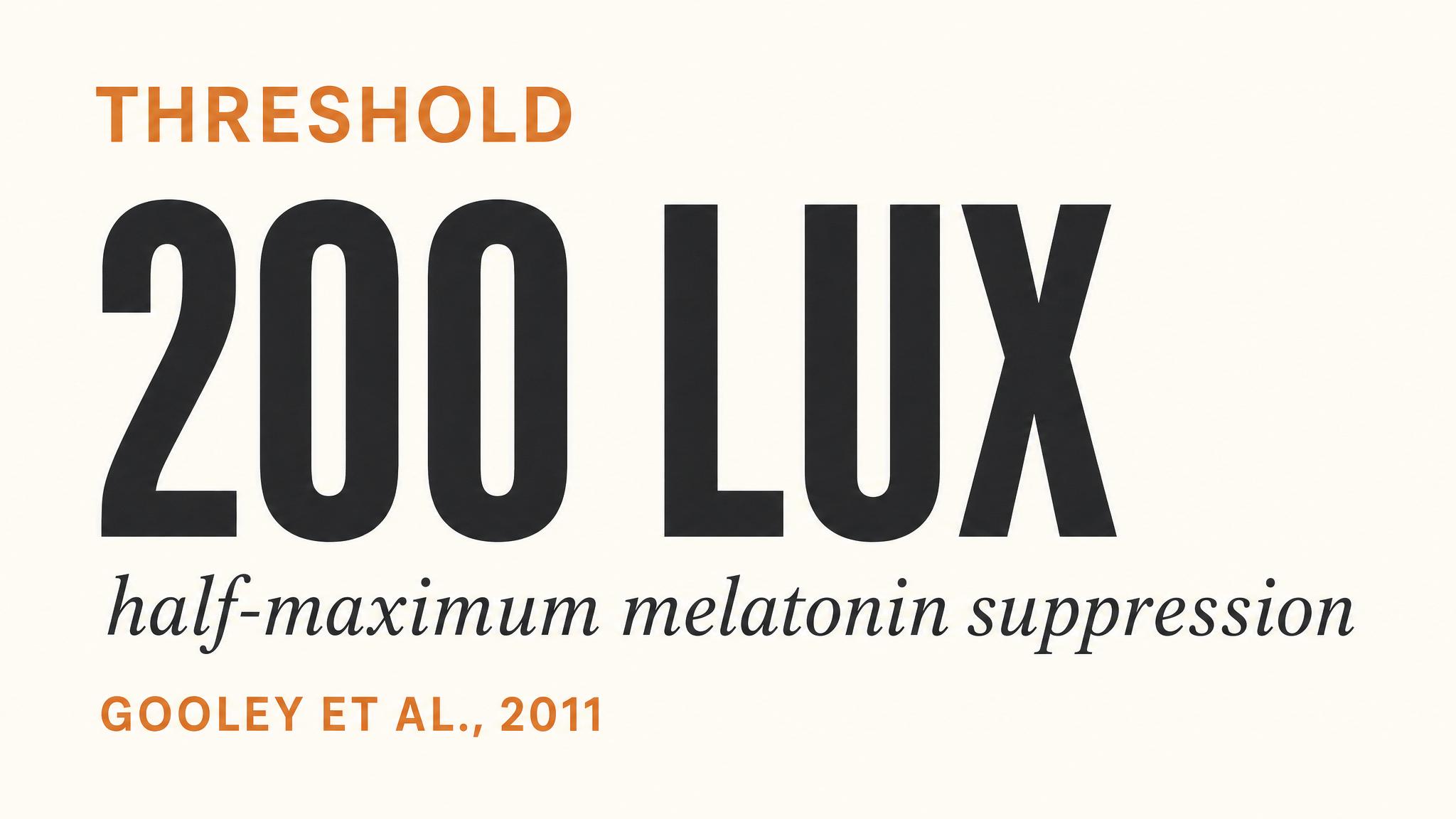







The Strategic Nap — Precision Rest in a Sleep-Deprived World
The final chapter synthesises the entire course by applying every concept students have learned to a single, practical question: how do you nap well? This is not a chapter about napping as a standalone topic — it is a…
Would you rather give up naps forever or give up coffee forever?
Impossible choice
Hot take: People who wake up refreshed from naps are lying. Every nap leaves you more tired and confused than before.
Unpopular opinion?
You have 30 minutes between classes. Do you nap or is that just asking for disaster?
Would you risk it?
Should your university have official nap pods on campus, or would you just never leave?
Quick poll
Someone told me to drink coffee then immediately nap. That's either genius or completely unhinged — which is it?
Genius or nonsense?
What's worse: pulling an all-nighter or taking an afternoon nap that wrecks your sleep schedule for three days?
Pick your poison
The Nap Calculator Duel
12-15 minutesInstructor presents 5 real-world scenarios on slides (e.g., 'It's 2pm, you have a presentation at 4pm, you got 5 hours of sleep last night'). Students turn to their neighbor and debate: Should they nap? For how long? Why? After 2 minutes of pair discussion per scenario, instructor cold-calls pairs to defend their choices. Class votes on most strategic answer using raised hands. Instructor reveals the science-backed optimal choice and awards points to winning pairs.
The 30-Minute Danger Zone Experience
8-10 minutesInstructor guides students through a visualization: 'Close your eyes. You've just woken from a 45-minute nap. Stand up if you'd feel...' followed by options (groggy/alert/confused). Students physically respond by standing/sitting. Instructor then describes the adenosine/sleep inertia mechanism while students are standing, making them literally 'feel' the stuck-in-between state. Pairs then share their worst nap-wake-up stories for 3 minutes, identifying which were likely in the danger zone. Instructor harvests 3-4 stories for whole-class analysis.
Coffee Nap Protocol Builder
15-18 minutesStudents work with their immediate neighbors (groups of 2-3) to design a 'Coffee Nap Protocol' for a specific persona (assigned by row: shift worker, new parent, grad student, athlete, etc.). Protocol must specify: coffee timing/amount, nap duration, environment setup, and expected outcomes. Groups write their protocol on paper. Instructor collects 6-8 protocols, displays them anonymously via document camera, and class critiques them using thumbs up/middle/down for adenosine timing accuracy. Top protocols get explained by their creators.
Circadian Window Live Mapping
10-12 minutesInstructor divides class into 4 sections. Each section represents a nap time window (12-1pm, 1-2pm, 2-3pm, 3-4pm). Students move to the section that matches when they typically feel most sleepy post-lunch. Once redistributed, students discuss with their new neighbors: What time do you actually nap? What duration? How does it affect your nighttime sleep? Each section nominates a spokesperson to report patterns. Instructor plots findings on board, revealing the class's collective circadian rhythm and discussing outliers/night shift workers.
The 90-Minute Gamble: Risk/Reward Analysis
10-12 minutesPresent the dilemma: '90-minute naps complete a sleep cycle and avoid inertia BUT risk disrupting nighttime sleep and require significant time.' Students position themselves on a stance spectrum by standing: far left = '90-min naps are worth it,' far right = '20-min naps only,' middle = 'depends on context.' From their seats, students debate with the person next to them who chose differently, defending their position with science concepts. After 4 minutes, instructor invites 3 students from different positions to present their best argument. Class revotes to see if anyone changed their mind.
Nap Myth Buster Speed Round
8-10 minutesInstructor rapid-fires common nap myths on slides ('Napping is for lazy people,' 'Any nap is better than no nap,' 'Morning naps are just as effective as afternoon ones'). For each myth, students discuss with their neighbor for 30 seconds: True, false, or complicated? Then vote as a class using hand signals (fist=false, open hand=true, thumbs sideways=complicated). Instructor reveals answer with 60-second scientific explanation. Keep score by row - which row has the best nap science literacy? Winning row gets to leave 2 minutes early (or gets first pick of exam review topics next week).
Transcript
It is 1:47pm on a Tuesday. You finished lunch thirty minutes ago and you are now staring at a paragraph you have read three times without absorbing a single word. Your eyelids are heavy. Your concentration has evaporated. You know, rationally, that you slept seven hours last night — not terrible — but something in your biology is pulling you toward unconsciousness with surprising force. You have forty-five minutes before your next obligation. Should you nap? And if so, for how long?
[short pause]
This question sounds simple. It is not. The difference between a nap that leaves you razor-sharp and one that leaves you disoriented and groggy for the next hour comes down to decisions about timing, duration, and context — decisions that draw on nearly every concept you have studied in this course. A well-designed nap is a tiny masterclass in sleep science. A poorly designed one is a case study in what happens when you ignore it. This final chapter brings everything together.
[short pause]
What Actually Happens When You Nap
To design a nap strategically, you first need to understand what your brain does during one. Not all naps are created equal, and their value depends entirely on which SLEEP STAGES you manage to reach — and which stage you are in when the alarm sounds.
Recall from Chapter Two that sleep unfolds in a predictable architectural sequence. When you close your eyes and drift off, you first enter Stage N-one, a brief transitional phase lasting only a few minutes. You then descend into Stage N-two, where the brain produces characteristic SLEEP SPINDLES — short bursts of oscillatory activity that play a critical role in memory consolidation, particularly for motor skills and declarative facts. If you remain asleep long enough, you sink further into Stage N-three, or slow-wave sleep, the deepest and most restorative phase. And if you sleep for roughly ninety minutes, you will typically complete a full cycle that includes REM sleep, the stage most strongly associated with creative problem-solving, emotional regulation, and the integration of new information with existing knowledge.
This architecture is not just academic trivia — it is the key to understanding why nap duration determines nap quality so dramatically.
[short pause]
The Power Nap: Ten to Twenty Minutes
A brief nap of ten to twenty minutes keeps you in N-one and N-two. You capture sleep spindles, enjoy a modest reduction in ADENOSINE — the homeostatic sleep pressure molecule you first encountered in Chapter One — and wake before the brain descends into the neurochemical quicksand of deep sleep. The result is a clean, crisp improvement in alertness and reaction time. The landmark NASA study by Rosekind and colleagues in nineteen ninety-five demonstrated this beautifully: long-haul pilots given a forty-minute nap opportunity, during which they achieved an average of twenty-six minutes of actual sleep, showed a fifty-four percent improvement in physiological alertness and a thirty-four percent improvement in performance compared to a no-rest control group. These pilots were not sleeping for hours — they were capturing N-one and N-two, and the payoff was immediate and substantial.
Similarly, Hayashi, Masuda, and Hori in two thousand three found that a twenty-minute afternoon nap was remarkably effective at restoring alertness against the mid-afternoon sleepiness trough, with effects that outperformed bright light exposure and face washing alone. The brevity is the point: you get in, you get the benefit, you get out before things get complicated.
[short pause]
The Full Cycle: Ninety Minutes
At the other end of the spectrum sits the ninety-minute nap — a complete sleep cycle that takes you through N-one, N-two, N-three, and back up through REM. As Mednick, Nakayama, and Stickgold demonstrated in two thousand three, naps of sixty to ninety minutes containing both slow-wave sleep and REM produced learning benefits on perceptual tasks that were statistically indistinguishable from a full night of sleep. Read that again: a single afternoon nap rivaled eight hours of nocturnal sleep for certain types of learning. The slow-wave component handled memory consolidation, and the REM component enabled creative integration and insight.
The ninety-minute nap is powerful. But it requires time, planning, and an understanding of what happens if you wake up at the wrong moment — which brings us to the concept that makes nap design genuinely tricky.
[short pause]
Sleep Inertia: The Danger Zone
SLEEP INERTIA is the grogginess, confusion, and impaired performance you experience immediately after waking from sleep — particularly from deep sleep. It is not just feeling a bit drowsy. Sleep inertia involves measurable decrements in cognitive function, reaction time, and decision-making that can persist for fifteen to thirty minutes or, in severe cases, over an hour. If you have ever woken from an afternoon nap feeling worse than before you lay down, you have experienced sleep inertia firsthand.
The critical variable is which sleep stage you are in when the alarm fires. Waking from N-one or N-two produces minimal inertia — you feel refreshed almost immediately. Waking from N-three, however, is like being pulled from the bottom of a deep pool. Your prefrontal cortex — the brain region responsible for executive function, judgment, and planning — is the last area to fully reboot after deep sleep arousal. This is why the thirty to sixty minute nap is often called the DANGER ZONE: it is long enough to descend into N-three but not long enough to complete the cycle and ascend back through lighter stages before waking.
According to Hilditch, Dorrian, and Banks in their comprehensive systematic review in two thousand seventeen, the pattern is clear. Naps shorter than approximately fifteen minutes rarely involved any slow-wave sleep and produced negligible inertia. Naps of around thirty minutes were unpredictable — some participants entered N-three, some did not — making this duration a gamble. And naps of sixty minutes almost always involved substantial N-three, meaning the sleeper was likely to be deep in slow-wave sleep when the alarm sounded. The review concluded that if your goal is a quick, clean alertness boost with minimal inertia risk, keep it under twenty minutes. If you have the luxury of time, extend to ninety minutes and allow the full cycle to complete naturally.
[short pause]
Before reading further, recall what you learned in Chapter Two about the function of N-three slow-wave sleep. Why would the brain resist being pulled out of this stage? What restorative processes are underway during N-three that might explain why interrupting them produces cognitive impairment?
[short pause]
The practical takeaway is stark: there is no such thing as a medium nap. You either nap short, under twenty minutes, or nap long, approximately ninety minutes. Anything in between risks placing you squarely in the danger zone — awake, technically, but cognitively impaired and wishing you had never closed your eyes.
[short pause]
The Coffee Nap: Engineering a Double Hit
Now that you understand nap architecture and sleep inertia, you are ready for one of the most elegant applications of sleep science: the COFFEE NAP, sometimes called the nappuccino. This strategy sounds almost too clever to work, but it exploits a precise pharmacological timing window that becomes obvious once you recall what you learned about caffeine in Chapter Four.
Here is the protocol: drink a cup of coffee — quickly, not sipped over twenty minutes — and immediately lie down for a twenty-minute nap. Then wake up.
Why does this work? Remember that caffeine does not give you energy. It blocks adenosine receptors, preventing the drowsiness signal from reaching your neurons. But caffeine takes approximately twenty to twenty-five minutes after ingestion to be absorbed through the small intestine, enter the bloodstream, and cross the blood-brain barrier in sufficient concentration to occupy those receptors effectively. During those twenty minutes, the caffeine is essentially in transit — it is not yet doing anything.
Meanwhile, your twenty-minute nap is doing something: sleep naturally clears adenosine. Not all of it — a brief nap only reduces adenosine modestly — but enough to partially lower the concentration of the very molecule that caffeine is about to start blocking. When you wake at the twenty-minute mark, two things happen simultaneously: the nap has reduced your adenosine levels, and the caffeine arrives at the receptors to block whatever adenosine remains. The result is a combined alertness boost that exceeds what either strategy produces alone.
As Hayashi and colleagues tested directly in two thousand three, caffeine consumed before a twenty-minute nap produced superior alertness restoration compared to a nap alone, caffeine alone, or other countermeasures like bright light exposure. The effect is not additive in a simple sense — it is synergistic, because the two mechanisms target the same system, adenosine signaling, from complementary angles.
[short pause]
From Chapter Four, recall the half-life of caffeine: approximately five to six hours for most adults. If you execute a coffee nap at two P M with one hundred milligrams of caffeine, approximately how much caffeine will still be active in your system at ten P M? How might this affect your nighttime sleep onset?
[short pause]
There is an important constraint here: the coffee nap only works if you actually fall asleep. If you are not sleepy enough to drift off quickly, or if you spend fifteen minutes lying awake anxiously watching the clock, you lose the adenosine-clearing benefit of the nap while still absorbing the caffeine. The strategy works best during the post-lunch dip — the very window when your biology is most cooperative about letting you fall asleep quickly. Which brings us to the question of timing.
[short pause]
Circadian Nap Timing: Why the Afternoon Window Matters
You now know how long to nap and what to drink beforehand. The remaining question is when. And the answer has been sitting in your notes since Chapter One.
Recall the TWO-PROCESS MODEL of sleep regulation: Process S, homeostatic sleep pressure driven by adenosine accumulation, rises steadily across waking hours, while Process C, the circadian alerting signal, oscillates on a roughly twenty-four-hour rhythm. During most of the day, Process C actively opposes Process S, keeping you alert even as adenosine accumulates. But there is a characteristic dip in the circadian alerting signal that typically occurs between approximately one P M and three P M — the POST-LUNCH DIP.
As Monk established in two thousand five, this afternoon trough in alertness and performance is fundamentally circadian in origin, not merely a consequence of having eaten lunch. People who skip lunch still experience the dip. It reflects a genuine secondary peak in sleep propensity — a brief window when Process C relaxes its opposition to Process S, and the accumulated adenosine from your morning hours gets a window of opportunity to express itself as drowsiness. Strijkstra and colleagues modeled this mathematically in two thousand three, demonstrating that the multiplicative interaction between homeostatic and circadian processes produces a characteristic double-peaked sleep propensity curve, with the secondary peak falling squarely in the early afternoon.
This is your optimal nap window. Napping during the post-lunch dip works with your biology rather than against it. You fall asleep faster, because both Process S and the circadian dip are cooperating. You are less likely to oversleep into problematic deep sleep during a short nap. And critically, you wake with enough remaining daylight hours to rebuild adequate sleep pressure before your nighttime sleep episode.
[short pause]
The Late Nap Problem
What happens if you nap at five or six P M instead? Here is where the course concepts converge in a cautionary way. A late-afternoon nap clears a significant portion of your accumulated adenosine just as your circadian system is preparing for its evening rise in alerting signal, before the subsequent decline toward the sleep gate around ten to eleven P M. The result: when bedtime arrives, you do not have enough homeostatic sleep pressure to fall asleep easily. You lie awake. Your sleep onset is delayed. Your total sleep time is reduced. And the next morning, you are more tired than you would have been had you simply endured the afternoon sleepiness and gone to bed on time.
Think of it this way: a late nap mimics the disruptive effect of evening light exposure, which you studied in Chapter Three. Evening light shifts your circadian clock later, delaying the sleep gate. A late nap does something analogous from the homeostatic side — it depletes Process S at precisely the wrong time, creating a mismatch between your circadian rhythm, which expects you to be sleepy, and your homeostatic drive, which has been partially reset by the nap. The effect is the same: you cannot fall asleep at your normal time.
Importantly, however, well-timed naps do not carry this risk. As Boukhris and colleagues recently demonstrated in two thousand twenty-five, both twenty-five-minute and ninety-minute afternoon naps, when completed by three P M, did not adversely affect subsequent nighttime sleep in student athletes. Total sleep time, sleep stage proportions, and wake-after-sleep-onset were all preserved. The key variable was not nap duration but nap timing: naps that ended within the post-lunch dip window left nighttime sleep architecture intact.
[short pause]
Consider your own chronotype. If you are an evening type whose natural circadian rhythm runs approximately two hours later than the average, how would you adjust the one P M to three P M nap window? What about a strong morning type who wakes naturally at five-thirty A M?
[short pause]
This interaction also explains a finding from Kurdziel, Duclos, and Spencer in two thousand twenty-one: multiple naps spread across a twenty-four-hour period cannot fully substitute for consolidated nighttime sleep. Slow-wave energy — a marker of deep restorative sleep — accumulated across six naps was significantly lower than what a single baseline night produced. Naps are supplements, not replacements. They work best as precision tools deployed at the right circadian moment, not as a strategy for avoiding proper sleep.
[short pause]
Putting It All Together: Your Personalised Nap Strategy
You have now assembled every piece of the puzzle. A strategic nap requires you to integrate knowledge from across this entire course:
Circadian timing from Chapter One: Nap during the post-lunch dip, adjusted for your chronotype.
Sleep architecture from Chapter Two: Choose either under twenty minutes, capturing N-one and N-two only, or approximately ninety minutes for a full cycle. Avoid the thirty to sixty minute danger zone.
Light management from Chapter Three: Dim your environment before napping to avoid alerting signals that inhibit sleep onset. After waking, seek bright light to accelerate the dissipation of sleep inertia.
Caffeine pharmacology from Chapter Four: If using a coffee nap, consume caffeine immediately before lying down. Ensure the dose and timing will not interfere with nighttime sleep. Account for the five to six hour half-life.
Temperature physiology from Chapter Five: A slightly cool environment facilitates sleep onset by supporting the core body temperature drop that accompanies the transition to sleep.
Arousal management from Chapter Six: Use a brief relaxation or cognitive deactivation technique to quiet the mind and accelerate sleep onset, maximizing actual sleep time within your nap window.
[short pause]
Each of these elements is a dial you can turn. Miss one — nap too late, choose the wrong duration, drink coffee at the wrong time, lie in a hot bright room while mentally rehearsing your to-do list — and the nap degrades from a precision tool into a source of frustration. Get them all right, and twenty minutes of afternoon rest can transform the cognitive quality of your entire remaining day.
[short pause]
A nap is not an escape from wakefulness. It is an investment in it — but only if you understand the terms of the investment.
[short pause]
Napping as Applied Sleep Science
There is something deeply satisfying about the fact that this course ends with napping — a behavior most people think of as simple, even lazy. But as you now understand, a well-designed nap is anything but simple. It requires an understanding of circadian biology, sleep-stage architecture, neurochemistry, pharmacology, thermoregulation, and psychological arousal management. It requires you to think about your body as a system with interacting processes operating on different timescales.
The broader lesson is this: sleep is not a passive state that happens to you. It is an active biological process that you can understand, respect, and — within limits — strategically manage. You cannot override your circadian rhythm any more than you can override gravity, but you can work with it. You cannot eliminate adenosine accumulation, but you can time its clearance intelligently. You cannot avoid sleep inertia if you wake from deep sleep, but you can avoid waking from deep sleep in the first place.
Throughout this course, you have built a conceptual toolkit — Process S and Process C, sleep spindles and slow waves, melanopsin and the S-C-N, adenosine receptors and caffeine half-lives, core body temperature curves, and the ascending arousal system. The strategic nap is simply the final exam for that toolkit: a single, practical problem that you can now solve with genuine understanding rather than guesswork.
Sleep well. And when the afternoon demands it, nap well too.
[short pause]
Key Takeaways
Short naps of ten to twenty minutes capture N-one and N-two sleep stages, providing alertness and memory benefits with minimal sleep inertia risk.
Full-cycle naps of approximately ninety minutes include N-three and REM, offering deep restoration and creative insight comparable to nocturnal sleep for some tasks.
The thirty to sixty minute range is the danger zone — long enough to enter deep N-three sleep but too short to complete the cycle, maximizing sleep inertia.
Coffee naps exploit the approximately twenty-minute caffeine absorption window: drink coffee, nap for twenty minutes, and wake as caffeine arrives at adenosine receptors already partially cleared by sleep.
The optimal nap window, typically one P M to three P M adjusted for chronotype, aligns with the post-lunch dip in the circadian alerting signal.
Late naps after approximately four P M reduce homeostatic sleep pressure at the wrong time, potentially disrupting nighttime sleep onset and architecture.
Well-timed naps completed by three P M do not adversely affect subsequent nighttime sleep quality or duration.
A strategic nap integrates knowledge of circadian timing, sleep architecture, light, caffeine, temperature, and arousal — the complete toolkit of this course.
[short pause]
Looking Ahead
This chapter concludes the course, but your understanding of sleep science is just beginning. Every night you sleep and every afternoon you feel the pull of drowsiness, you now have a framework for understanding what your body is doing and why. The concepts from these seven chapters — circadian rhythms, sleep architecture, light sensitivity, caffeine dynamics, thermoregulation, arousal systems, and strategic napping — are not isolated facts. They are an integrated model of one of the most fundamental processes in human biology. Take it with you. Use it. Sleep is not a luxury to be minimized — it is the foundation on which every waking hour is built.